

2. Блокування біосинтезу білка дифтерійним токсином (адф-рибозилювання факторів трансляції)
П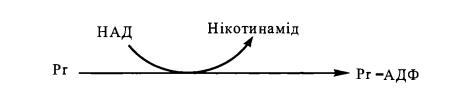 ри
АДФ-рибозилуванні відбувається приєднання
до білка, що підлягає ковалентній
модифікації, АДФ-рибозильного радикалу,
який відщеплюється від складної молекули
нікотинамідаденіндинуклеотиду (НАД)
ри
АДФ-рибозилуванні відбувається приєднання
до білка, що підлягає ковалентній
модифікації, АДФ-рибозильного радикалу,
який відщеплюється від складної молекули
нікотинамідаденіндинуклеотиду (НАД)
АДФ-рибозилування білків є механізмом регуляції багатьох видів клітинної активності, зокрема, експресії ядерного генетичного апарату. Особливим прикладом біологічного ефекту такої модифікації ферментного білка є дія холерного токсину, каталітична субодиниця якого АДФ-рибозилує аденілатциклазу мембран ентероцитів. Це, у свою чергу, супроводжується стимуляцією фосфорилування та активацією хлорного каналу мембран, який є відповідальним за транспорт у просвіт кишківника іонів СІ" та води, що і призводить до важкої діареї - головного клінічного прояву холери.
3. Джерело енергії для м’язового скорочення. Атф, креатинфорфат. Особливості обміну в серцевому м’язі.
м'язове скорочення, в основі якого лежить переміщення товстих і тонких філаментів, потребує участі АТФ; циклічне перетворення АТФ в АДФ є необхідною передумовою як скорочення, так і
розслаблення м'язів. Гідроліз АТФ до АДФ та Фн здійснюється завдяки АТФ-азній активності
глобулярних головок S1 міозину.
A. У м'язі, що перебуває в стані спокою, S1-головки міозину не сполучені з актиновими філамен-тами. Продукти гідролізу АТФ (АДФ та Фн) зв'язані з міозином.
B. При збудженні м'яза S1-головки зсуваються в напрямку тонких філаментів і сполучаються з
нитками актину (G-субодиницями). Фн вивільняється з комплексу з міозином.
C. Вивільнення АДФ з комплексу з міозином супроводжується конформаційним зсувом у
просторовому розташуванні головки S1, що зв'язана з актином (зміщенням кута між головкою і віссю міофібрили з 90° на 45°).
D. Взаємодія з актином молекули АТФ супроводжується розривом зв'язку між актином і міозином. S1-головка знову віддаляється від тонкого філамента.
E. АТФ, що вивільнився, гідролізується до АДФ та Фн, завдяки АТФ-азній активності вільних головок міозину. Продукти гідролізу знову сполучаються з міозином. Актинові та міозинові філамента готові до нового циклу взаємодії та пересування.

Генерація АТФ із креатинфосфату є найбільш швидким механізмом утворення АТФ при потребі термінового включення процесу м'язового скорочення. За рахунок дії креатинфосфокіназного (КФК) механізму можливе забезпечення інтенсивної роботи м'язів протягом 2-5 с - періоду, необхідного для підключення інших біоенергетичних механізмів. У період розслаблення м'язів відбувається ресинтез креатинфосфату за рахунок зворотної течії креатинкіназної реакції.
Міокард за своєю структурою та біохімічними властивостями близький до смугастих м'язів, зокрема до червоної скелетної мускулатури. Особливістю енергетичного обміну в міокарді є майже повністю аеробний характер, що зумовлює його високу чутливість до порушень у постачанні кисню, зокрема внаслідок звужень у коронарних артеріях. Як субстрат біологічного окислення міокард використовує здебільшого жирні кислоти, на окислення яких витрачається близько 70 % 02, що споживає серцевий м'яз (А. Я. Николаев, 1998). Частка жирних кислот у забезпеченні міокарда АТФ дещо зменшується після споживання їжі, а також в умовах фізичного навантаження; в зазначених умовах зростає окислення глюкози та молочної кислоти, відповідно.Регуляція м'язового скорочення в міокарді відбувається за механізмами, близькими до скелетних м'язів. Проте, на відміну від скелетних м'язів, головним джерелом Са2+ для скорочення кардіоміцитів є екстрацелюлярний кальцій, який при збудженні міокарда надходить у саркоплазму через канали плазматичних мембран.