Класифікація гормонів: за місцем синтезу, хімічною природою, механізмом дії.
Гормони - фізіологічно активні сполуки (ФАС), що продукуються залозами внутрішньої секреції (ендокринними залозами) або іншими спеціалізованими клітинами і діють як сигнальні молекули та біорегулятори метаболічних процесів та фізіологічних функцій в організмі. Біологічні ефекти гормонів здійснюються у надзвичайно низьких концентраціях. Гормони, що синтезуються в ендокринних залозах ("справжні", істинні гормони), секретуються в кров'яне русло
і після перенесення спеціалізованими транспортними білками здійснюють свої біологічні ефекти, як правило, на відстані, тобто діють на віддалений чутливий орган або органи.
До "справжніх" гормонів належать:
- гормони гіпоталамуса та гіпофіза;
- гормони щитовидної залози;
- гормони паращитовидної залози;
-гормони ендокринних клітин підшлункової залози;
-гормони коркової частини надниркових залоз;
- гормони чоловічих та жіночих статевих залоз;
- гормони епіфіза.
Хімічна структура гормонів За хімічною будовою всі гормони поділяють на такі класи:
(1) білково-пептидні гормони (прості білки; глікопротеїни; пептиди): гіпоталамо-гіпофізарні гормони; гормони паращитовидної залози; гормони острівкової частини підшлункової залози; гастроінтестинальні гормони; нейропептиди; численні тканинні біорегулятори пептидної природи;
(2) гормони - похідні амінокислот: гормони щитовидної залози; гормоноїди мозкової частини надниркових залоз (катехоламіни); інші нейромедіатори з властивостями гістогормонів (серотонін, дофамін, гістамін); гормон епіфіза - мелатонін;
(3) гормони стероїдної природи: глюкокортикоїди та мінералокортикоїди кори надниркових залоз; чоловічі та жіночі статеві гормони; похідні вітаміну D;
(4) біорегулятори - похідні арахідонової кислоти (ейкозаноїди): простагландини, простацикліни, тромбоксани, лейкотрієни, ліпоксини.
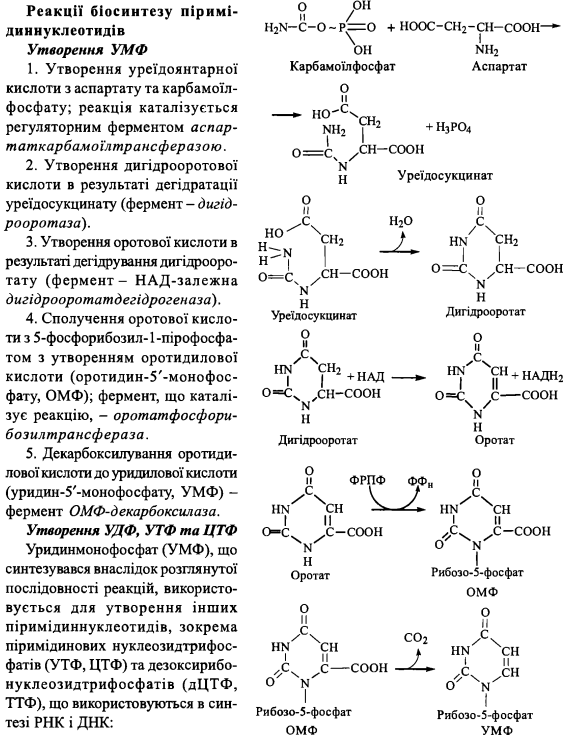
Біосинтез піримідинових нуклеотидів

Білет 7
Жіночі статеві гормони. Механізм дії
До жіночих статевих гормонів належать:
естрогени - похідні естрану (С18-стероїди);
прогестагени - похідні прегнану (С2І-стеро'їди).
Діють естрогени шляхом зміни синтезу білків у клітині, хоча можуть надавати і зовнішньорецепторний ефект. Механізм дії. Е. вибірково накопичуються в органах-мішенях — матці, піхві, молочних залозах, передній частці гіпофіза, печінці, де зв’язуються зі специфічним позаядерним білком естрофіліном, рецепторами плазматичних мембран клітин-мішеней, утворюючи з ними гормон-рецепторні комплекси. Вони проникають у ядро, активують синтез ДНК і РНК, впливають на синтез білка.
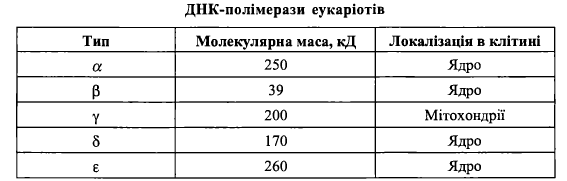
Загальна схема біосинтезу днк. Ферменти реплікації днк у прокаріотів та еукаріотів.
Було встановлено, що в цих умовах синтезується певна кількість нової ДНК (точніше, полідезоксирибонуклеотиду), до складу якого входили дезоксинуклео- зидмонофосфати (дНМФ) із складу дНТФ, що були використані для біосинтезу. При цьому новий ланцюг ДНК утворювався за рахунок приєднання дНМФ до кінця одного з ланцюгів ДНК, що вже існувала (передіснуючої ДНК). Особливості синтезу ДНК в ДНК-полімеразній системі:
- ДНК-полімераза не може синтезувати повністю новий ланцюг ДНК, а спроможна лише приєднувати дНМФ до вже існуючого ланцюга;
- синтез нового ланцюга ДНК відбувається в напрямку 5'-3', тобто ДНК-поліме- раза послідовно приєднує нуклеотиди (дНМФ з наявних дНТФ) до 3'-кінця одного з ланцюгів ДНК ("ланцюга-затравки");
- для синтезу нового ланцюга ДНК необхідний також ланцюг-матриця; нуклеотиди сполучаються з З'-кінцем "ланцюга-затравки" відповідно до нуклеотидної послідовності в "ланцюгу-матриці" (тобто, за принципом комплементарності).
Ферменти біосинтезу ДНК у прокаріотів:
1) ДНК-полімераза І - білок з м.м. 103 кДа. Біохімічні функції:
5'—>3' - полімеразна активність (розглянута вище);
5'—>3' - екзонуклеазна активність (тобто спроможність видаляти нуклеотиди вище від напрямку синтезу);
З'—»5' - екзонуклеазна активність ("коригуюча" активність, або спроможність видаляти вже включений нуклеотид, якщо він включений помилково, тобто не є комплементарним ланцюга-матриці);
2) ДНК-полімераза II (білок з м.м. 120 кДа) не є основним в реплікації ДНК у прокаріотів; за існуючими уявленнями, переважною функцією цього ферменту є участь у репарації ДНК;
3) ДНК-полімераза III- головний фермент, що реалізує процес елонгації ДНК у Е. соїі.