

Nikolls_-_Ot_neyrona_k_mozgu
.pdfГлава 17. Трансдукция механических и химических стимулов |
363 |
Рис. 17.1. Короткие и длинные сенсорные рецепторные клетки. (А) Короткие рецепторы сетчатки, такие как палочки и колбочки, а также механочувствительные волосковые клетки имеют длину не более 100 мкм. (В) Длинные рецепторы, такие как афференты мышечных веретен и кожные механорецепторы, используют потенциалы действия для передачи своих сигналов на удаленный вторичный нейрон.
Fig. 17.1. Short and Long Sensory
Receptor Cells. (A) Short receptors
— such as retinal rods and cones, and mechanosensory hair cells of the inner ear — are less than 100 µπι in length. Thus, receptor potentials generated within the sensitive ending spread effectively throughout the cell, altering transmitter release at synaptic areas.
(B) Long receptors, such as muscle spindle afférents and cutaneous mechanoreceptors, employ action potentials to conduct their signals to a distant second-order neuron. The amplitude of the receptor potential is encoded in the frequency of action potentials it generates.
ны, а следовательно, и константа длины распространения пассивной деполяризации, необычайно высоки. Хотя рецепторные потенциалы обычно являются деполяризационными, некоторые короткие рецепторы реагируют на свои адекватные раздражители гиперполяризационным изменением потенциала. Это происходит, к примеру, в фоторецепторах сетчатки позвоночных (глава 19) и в кохлеарных волосковых клетках, в которых возникают как гиперполяризационные, так и деполяризационные ответы. Независимо от того, какова полярность рецепторного потенциала, короткие рецепторы тонически высвобождают нейромедиатор из своих синаптических зон; при этом деполяризация усиливает, а гиперполяризация снижает исходный уровень высвобождения.
В длинных рецепторах (рис. 17.1В), таких как рецепторы кожи или мышц, информация от одиночного рецептора может посылаться на гораздо большие расстояния, чтобы достичь сенсорных клеток второго порядка (например, от большого пальца ноги к спинному мозгу). Чтобы обеспечить это, рецептор совершает второе преобразование: рецепторный потенциал дает начало последовательности потенциалов действия, длительность и частота которой кодируют информацию о длительности и интенсивности исходного стимула. Затем этот импульсный разряд переносит данную информацию к синаптическим окончаниям клетки.
Частотное кодирование интенсивности стимула обеспечивается путем взаимодействия между стационарным рецепторным током от сенсорных окончаний и сдвигами проводимости во время потенциала действия. Повышение калиевой проводимости, происходящее во время фазы восстановления в конце каждого потенциала действия, гиперполяризует мембрану, смещая мембранный потенциал в направлении ЕК (калиевого равновесного потенциала). Это увеличение калиевой проводимости, однако, весьма кратковременно, в результате чего поддерживаемый за счет постоянного растяжения трансдукционный ток вновь деполяризует мембрану до уровня генерации импульса. Чем сильнее рецепторный ток, тем скорее вновь достигается пороговый уровень генерации разряда и тем выше становится частота импульсации. Подобный анализ применим также ко всем нейронам, в которых любой синаптический вход, по аналогии

364 |
Раздел III. Интегративные механизмы |
Рис. |
17.2. |
Внеклеточно |
регистрируемые |
рецепторные |
|
потенциалы |
от |
|
чувствительных |
нервных |
|
волокон |
|
мышечного |
веретена. (А) Растяжение мышцы (верхняя кривая) вызывает рецепторный потенциал, сопровождающийся серией потенциалов действия (нижняя кривая). (В) Четыре уровня растяжения мышцы после добавления прока ина, блокирующего потенциалы действия (кроме первого). (С) Зависимость между амплитудой рецепторного потенциала и увеличением длины мышцы.
Fig. 17.2. Receptor Potentials Recorded Extracellularly from a sensory nerve fiber supplying a muscle spindle. The recording electrode is placed as close as possible to the receptor. Downward deflection of the voltage
record (tower traces) indicates receptor depolarization. (A) Stretching the muscle (upper trace) produces a receptor potential, upon which is superimposed a series of action potentials (lower trace). (B) Four stretches of increasing magnitude applied to the muscle after procaine has been added to the bathing solution. Action potentials (except for the first) are abolished by procaine, but the receptor potentials remain.
(C) Plot of receptor potential amplitude against increase in muscle length. (After Katz, 1950.)
с рецепторным потенциалом, влияет на изменение частоты потенциалов действия.
Кодирование параметров стимула рецепторами растяжения
То, каким образом сенсорные рецепторы генерируют электрические сигналы, довольно давно было показано Эдрианом и Зоттерманом7) с использованием внеклеточного отведения от сенсорных нервных волокон, идущих от мышечных рецепторов растяжения (мышечных веретен) позвоночных. Первая демонстрация связи между сенсорными стимулами и электрическими сигналами в механорецепторе была получена Катцем8), который, регистрируя рецепторные потенциалы, показал, что растяжение вызывает деполяризацию сенсорного окончания. Когда рецепторный потенциал удалось зарегистрировать изолированно, путем блокады нервного разряда местным анестетиком прокаином, стало очевидно, что амплитуда этого потенциала растет как степенная функция от величины мышечного растяжения.
Рис. 17.2 показывает зависимость между амплитудой рецепторного потенциала и натяжением. В начале растяжения эта функция растет примерно на 0,1 мВ на миллиметр, но при более сильном растяжении скорость роста снижается. Чувствительность сенсорного окончания, выраженная в милливольтах на миллиметр растяжения, снижается по мере роста стимула. Многие сенсорные рецепторы используют преимущества этой нелинейной зависимости для того, чтобы обеспечить амплитудное кодирование в широком диапазоне интенсивности стимула. В таких рецепторах амплитуда ответа продолжает расти, но уже пропорционально логарифму интенсивности стимула. Это чрезвычайно полезно для таких рецепторов, как волосковые клетки и фоторецепторы, которые реагируют на стимулы, варьирующие по амплитуде на несколько порядков.
Зависимость между интенсивностью стимула и силой возникающих в ответ на него ощущений впервые описана в 1846 году Вебером. Он определял способность различать, какая из двух гирек тяжелее, держа их в разных руках, и показал, что способность к различению зависит от исходной величины гирек. То есть испытуемые были способны легко определить разницу в 3 грамма между двумя гирями, каждая из которых весила примерно по 100 грамм, тогда как килограммовые гири должны были отличаться на 30 грамм (в каждом случае различимая разница должна была быть около 3 % от веса каждого объекта). Эта зависимость была форма-

Глава 17. Трансдукция механических и химических стимулов |
365 |
Рис. 17.3. Рецептор растяжения ракообразных. (А) Живой рецепторный нейрон при освещении в темном поле. (В) Схема связи между рецептором растяжения и мышцей при внутриклеточном отведении.
Fig. 17.3. Crustacean Stretch Receptor. (A) Living receptor neuron viewed with dark-field illumination. Distal portions of six dendrites insert into the receptor muscle, which is not visible. (B) Relation between stretch receptor neuron and muscle, indicating the method of intracellular recording. The excitatory fiber to the muscle produces contraction; the inhibitory fiber innervates the neuron. Two additional inhibitory fibers are not shown. (After Eyzaguirre and Kuffler, 1955.)
лизована Фехнером, который пришел к заключению, что такой результат предполагает логарифмическую зависимость между стимулом и ответной реакцией. Закон Вебера— Фехнера 9) — это выражение нелинейной зависимости между силой раздражителя и ощущением. Хотя точная формула соотношения зависит от модальности раздражителя, эта зависимость применима ко многим аспектам восприятия и поведения. (Например, мы можем экономить копейки, прицениваясь к карандашам, но с удовольствием переплатим лишние дветри сотни за компьютер более высокого класса.)
Рецепторы растяжения речного рака
Кодирование стимула было детально проанализировано на примере рецепторов растяжения рака Эйзагиром и Куффлером 10). Этот препарат особенно удобен тем, что клеточное тело рецептора растяжения находится не в ганглии, а расположено изолированно на периферии, где его можно видеть в живом препарате (рис. 17.3А). Клетка достаточно велика для введения внутриклеточных микроэлектродов. Она проникает своими дендритами в ближайший тонкий мышечный тяж, а свой аксон посылает центрипетально в сегментарный ганглий (рис. 17.3В). В дополнение к этим связям, рецепторный нейрон получает тормозную иннервацию из ганглия; мышечное волокно, связанное с рецептором, в свою очередь, получает возбуждающую и тормозную эфферентную иннервацию. Таким образом, чувствительность рецептора регулируется ЦНС.
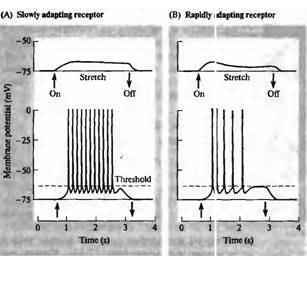
У ракообразных существует два типа рецепторов растяжения, различающихся своими структурными и физиологическими свойствами, а также тем, что они своими дендритами иннервируют различные типы мышц. Одни из них бурно активируются в начале растяжения, но затем их реакция быстро ослабевает. Такое снижение ответной реакции в процессе действия стимула постоянной интенсивности называется адаптацией. В противоположность быстро адаптирующемуся рецептору, второй тип называется медленно адаптирующимся; это означает, что его реакция сохраняется неизменной во время продолжительного растяжения. Реакции быстро адаптирующихся и медленно адаптирующихся рецепторов растяжения показаны на рис. 17.4. У медленно адаптирующихся рецепторов (рис. 17.4А) умеренное растяжение мышцы вызывает деполяризационный рецепторный потенциал амплитудой около 5 мВ, сохраняющийся неизменным во время растяжения. Более сильное растяжение приводит к увеличению потенциала, в результате чего
366 |
|
|
|
|
|
Раздел III. Интегративные механизмы |
||
Рис. |
17.4. |
Ответы |
нейронов |
|
|
|
||
рецептора |
растяжения |
на |
|
|
|
|||
увеличение |
длины |
мышцы |
при |
|
|
|
||
внутриклеточном |
|
отведении, |
|
|
|
|||
аналогичном рис. 17.3В. (А) В |
|
|
|
|||||
медленно |
адаптирующемся |
|
|
|
||||
рецепторе слабое |
растяжение в |
|
|
|
||||
течение |
2 |
секунд |
вызывает |
|
|
|
||
подпороговый |
|
рецепторный |
|
|
|
|||
потенциал. |
При |
|
усилении |
|
|
|
||
растяжения |
возникает |
серия |
|
|
|
|||
потенциалов действия. (В) В быстро |
|
|
|
|||||
адаптирующемся |
|
рецепторе |
|
|
|
|||
рецепторный |
потенциал снижается, |
|
|
|
||||
а при более сильном растяжении |
|
|
|
|||||
снижается |
частота |
|
потенциалов |
|
|
|
||
действия. |
|
|
|
|
|
|
|
|
Fig. 17.4. |
Responses |
of |
Stretch |
|
|
|
||
Receptor Neurons to increases in |
|
|
|
|||||
muscle |
length, recorded |
intracellularly |
|
|
|
|||
as indicated in Figure 17.3B. (A) In a |
|
|
|
|||||
slowly adapting receptor, a weak |
|
|
|
|||||
stretch for about 2 s produces a |
|
|
|
|||||
|
|
|
||||||
subthreshold |
receptor |
potential that |
|
up a series of action potentials (lower record). (B) In a rapidly adapting |
|
|||
persists through- out the stretch (upper |
|
receptor, the receptor potential is not maintained (upper record), and |
||||||
record). With a stronger stretch, a |
|
during the large stretch the action potential frequency declines (lower |
||||||
larger receptor potential sets |
|
|
record). (After Eyzaguirre and Kuffler, 1955.) |
|||||
клетка деполяризуется до порогового уровня, и в ней возникает серия потенциалов действия, которые распространяются центрипетально по аксону. В быстро адаптирующемся рецепторе подобное растяжение мышцы вызывает только кратковременные ответы (рис. 17.4В).
Мышечные веретена
Рецепторы растяжения в скелетных мышцах у млекопитающих демонстрируют те же механизмы действия, что и у ракообразных. Такие рецепторы растяжения в свое время были названы анатомами мышечными веретенами, поскольку они по форме напоминают веретена, используемые ткачами. Особенности того, как реагируют мышечные веретена, были исследованы Мэтьюзом в начале 1930-х годов11)--13). На протяжении многих лет его опыты оставались одним из лучших примеров всестороннего описания периферического органа чувств и механизмов управления его работой. Мэтьюзу удалось записать импульсы в одиночных нервных волокнах от отдельных веретен лягушек и кошек с помощью осциллоскопа, который он сконструировал для этой цели (в 1930 году это было настоящим подвигом).
Мышечные волокна в составе веретена (интрафузальные волокна) отличаются от основной мышечной массы (экстрафузальных волокон) во многих отношениях (см. обзор Ханта14)), включая молекулярную структуру содержащегося в них миозина15). Они названы интрафузальными волокнами, от лат. fusus — веретено. Рис. 17.5 схематически иллюстрирует сенсорный аппарат веретен в мышцах конечности кошки. Веретено состоит из капсулы, содержащей от 8 до 10 интрафузальных волокон. В центральной, или экваториальной, области каждого волокна находится большое скопление ядер. Их расположение дает основание для классификации интрафузальных волокон, которые подразделяют на сумчатые и цепочечные волокна (bag or chain fibers), в зависимости от того, сгруппированы ли ядра в центре или распределены линейно вдоль волокна.
Два типа сенсорных нейронов иннервируют каждое мышечное веретено. Более крупные нервные волокна, афференты группы la, имеют диаметр от 12 до 20 мкм и проводят импульсы со скоростью до 120 м/с. (Классификация волокон, которая используется здесь и является общепринятой, приведена в главе 7.) Их терминали обвиваются вокруг центральной части как сумчатых, так и цепочечных волокон, и формируют первичные окончания. Более мелкие сенсорные нервы (волокна группы II) имеют диаметр от 4 до 12 мкм и проводят импульсы гораздо медленнее. Они контактируют с цепочечными волокнами, где образуют вторичные окончания.

Глава 17. Трансдукция механических и химических стимулов |
367 |
Рис. 17.5. Мышечное веретено млекопитающего. (А) Схема иннервации веретена. (В) Упрощенная диаграмма типов интрафузальных мышц и их иннервации.
Fig. 17.5.Mammalian Muscle Spindle. (A) Scheme of mammalian muscle spindle innervation. The spindle, composed of small intrafusal fibers, is embedded in the bulk of the muscle, which is made up of large muscle fibers supplied by a motoneurons. 7 Motor (fusimotor) fibers supply the intrafusal muscle fibers, and group I and group II afferent fibers carry sensory signals from the muscle spindle to the spinal cord.
(B) Simplified diagram of intrafusal muscle types and their innervation. (В after Matthews, 1964.)
Мышечное веретено иннервируется, кроме того, мотонейронами (фузимоторные волокна, или γ-мотонейроны). Они обеспечивают сокращение интрафузальных волокон и растягивают за счет этого центральную ядерную область, где расположены сенсорные окончания, вызывая в них разряд импульсов. Это взаимодействие обеспечивает механизм эфферентного контроля чувствительности мышечного веретена, который будет описан в главе 22.
Реакция на статическое и динамическое мышечное растяжение
Когда мышца, а следовательно, и находящиеся в ней веретена подвергаются быстрому растяжению, то в сенсорных волокнах групп Ia и II возникают рецепторные потенциалы и разряды импульсов. Характеристики разрядов в двух типах окончаний, однако, существенно отличаются. Первичные окончания, связанные с более крупными аксонами группы Ia, чувствительны в основном к изменениям степени растяжения. Поэтому частота разрядов в них максимальна во время динамической фазы, пока растяжение возрастает, а затем поддерживается на более низком стабильном уровне до тех пор, пока растяжение сохраняется. Вторичные окончания, связанные с более мелкими волокнами группы II, относительно слабо реагируют на изменения степени растяжения, но чувствительны к уровню статического напряжения. Такое поведение иллюстрируется рис. 17.6. Группа Ia (динамическая) и группа II (статическая) являются, соответственно, аналогами быстро адаптирующихся и медленно адаптирующихся рецепторов в мышце рака и в других сенсорных системах.
Механизмы адаптации в механорецепторах
В мышечных веретенах млекопитающих постепенное снижение деформации сенсорных окончаний обеспечивается за счет упруго--эластических свойств интрафузальных воло-

368 |
Раздел III. Интегративные механизмы |
Рис. 17.6. Различия ответов мышечных веретен. Записи потенциалов действия от одиночного первичного и вторичного сенсорных афферентных волокон.
Fig. 17.6. Differences in Muscle Spindle Responses. Recordings of action potentials from single primary (group la) and secondary (group II) sensory afferent fibers originating in a cat muscle spindle. The primary fiber greatly increases its discharge rate as tension develops during the stretch; during the maintained phase of the stretch, it quickly adapts to a lower rate. The secondary fiber increases its firing rate more slowly as tension develops and maintains its discharge during the steady stretch. (After Jansen and Matthews,
1962.)
кон17). В рецепторах растяжения ракообразных также обнаружены разнообразные процессы, способствующие адаптации16· 18) -- 20). В медленно адаптирующемся рецепторе растяжения пачечная импульсная активность приводит к повышению внутренней концентрации натрия и к активации натрий-калиевого насоса. Создаваемый им суммарный перенос положительных зарядов из клетки во внеклеточную среду снижает амплитуду рецепторного потенциала, а вследствие этого и частоту разрядов. Еще один фактор, способствующий адаптации, — это увеличение калиевой проводимости. Например, во время серии импульсов в рецепторе растяжения рака поступление кальция через потенциалзависимые каналы вызывает открывание кальций-активируемых калиевых каналов. Результатом этого повышения калиевой проводимости является «сокращение» рецепторного потенциала, проявляющееся в уменьшении его амплитуды и частоты сенсорных импульсов.
Быстро адаптирующийся рецептор растяжения рака незамедлительно реагирует адаптацией частоты импульсации даже в ответ на экспериментально приложенный постоянный деполяризующий ток. В процессе принудительного растяжения кальций, входящий через трансдукционные каналы, активирует близлежащие кальций-зависимые калиевые каналы, обеспечивая, тем самым, гиперполяризацию клетки21).
Адаптация в тельце Пачини
Тельце Пачини — это быстро адаптирующийся кожный механорецептор22). Его нервное окончание заключено в капсулу, напоминающую по форме луковицу. Если подвергать капсулу медленному сдавливанию, то никакого ответа не возникает; более быстрое сдавливание вызывает только один или два потенциала действия. Однако эти рецепторы чрезвычайно чувствительны к вибрации на частотах до 1 000/с. Хотя их обнаруживают повсеместно в подкожных тканях, особенно часто они встречаются вокруг подушечек лап и возле когтей млекопитающих, а также в межкостных связках, соединяющих кости в передних и задних конечностях, где они действуют как чувствительный детектор колебаний почвы 23). Сходное образование, тельце Хербста, обнаружено у птиц в ногах, клювах и в кожной ткани (а у дятлов
— даже в языке!). В качестве примеров выполняемых ими физиологических функций можно привести восприятие уткой с помощью клюва колебаний воды, вызываемых ее мелкой добычей, и определение парящей птицей вибраций оперения, возникающих в полете из-за незначительных аэродинамических отклонений24).
Механизм адаптации в тельце Пачини был детально изучен Вернером Левенштейном и его сотрудниками, которые показали, что это происходит отчасти благодаря динамическим свойствам капсулы25). Когда на изолированную интактную капсулу наносили пульсирующее механическое воздействие, то короткий рецепторный потенциал возникал в начале и в конце каждого биения (рис. 17.7А). Ответы на постоянное давление столь кратковременны, поскольку сжатие чувствительного окончания ослабляется благодаря перераспределению жидкости в капсуле. После того как капсулу осторожно удалили с нервного окончания, рецепторный потенциал начал медленно спадать во время пульсового сти-

Глава 17. Трансдукция механических и химических стимулов |
369 |
||||||||
Рис. 17.7. |
Адаптация |
в |
|
|
|||||
тельце Пачини. (А) Скачок |
|
|
|||||||
давления, |
приложенный |
|
|
||||||
к |
тельцу, |
вызывает |
|
|
|||||
быстро |
адаптирующийся |
|
|
||||||
рецепторный |
потенциал. |
|
|
||||||
Аналогичный |
|
|
ответ |
|
|
||||
возникает |
|
в |
момент |
|
|
||||
снятия давления. (В) |
|
|
|||||||
После |
удаления |
капсулы |
|
|
|||||
давление, |
приложенное |
к |
|
|
|||||
нервной |
|
терминали, |
|
|
|||||
вызывает |
|
рецепторный |
|
|
|||||
потенциал, |
|
длящийся |
|
|
|||||
до |
конца раздражения. |
|
|
|
|||||
Fig. 17.7. Adaptation in a |
|
|
|||||||
Pacinian Corpuscle. |
(A) |
A |
|
|
|||||
pressure step applied to the |
|
|
|||||||
body of the corpuscle (lower |
|
|
|||||||
trace) |
produces |
a |
rapidly |
|
|
||||
adapting receptor |
potential |
|
|
||||||
(upper trace), as a result of |
the capsule to the nerve terminal. A similar response occurs on removal of the pulse. (B) |
||||||||
a |
transient |
wave |
of |
After removal of the capsule layers, pressure applied to the nerve terminal produces a |
|||||
deformation |
|
that |
|
travels |
receptor potential that lasts for the duration of |
the pulse. (After Loewenstein and |
|||
through |
|
|
|
|
|
|
Mendelson, 1965.) |
|
|
мула (рис. 17.7В). Тем не менее, даже когда рецепторный потенциал был пролонгирован, в афферентном аксоне все равно возникала лишь короткая вспышка потенциалов действия (на рисунке не показано); это означает, что свойства аксона как такового совпадают со свойствами интактного рецептора.
§ 2. Трансдукция механических стимулов
Очевидно, что должны существовать чувствительные к растяжению ионные каналы, расположенные на механочувствительных нервных окончаниях, где генерируется рецепторный потенциал. Более того, эти каналы трансдукции должны отличаться от каналов, обеспечивающих генерацию потенциала действия, поскольку ответы на растяжение сохраняются в присутствии местного анестетика, который блокирует распространение возбуждения. Механочувствительные ионные каналы обнаружены в самых разнообразных клетках и органах, включая эндотелиальные клетки кровеносных сосудов, барорецепторы в каротидном синусе, рецепторы прикосновения и давления в коже, мышечные рецепторы растяжения и Механочувствительные волосковые клетки внутреннего уха26)--28).
Методом фиксации напряжения было показано, что токи, лежащие в основе рецепторного потенциала в рецепторах растяжения рака, обусловлены увеличением проницаемости как для натрия и калия29· 30), так и для двухвалентных катионов31), а также для более крупных органических катионов, таких как трис (три[гидроксиметил]аминометан) и аргинин. Увеличение проводимости, вызванное растяжением, не чувствительно к тетродотоксину32), однако подвержено влиянию некоторых местных анестетиков33). Рецепторные потенциалы в мышечных веретенах позвоночных также связаны с увеличением катионной проницаемости34).
Одиночные каналы, активируемые прогибанием мембраны (membrane distortion), впервые наблюдали в мембранных кусочках (пэтчах) из эмбриональных мышечных клеток цыпленка35) и в мембранах других клеток, не имеющих отношения к сенсорной трансдукции36). Пэтчотведения активности похожих каналов, сделанные на первичных дендритах рецептора растяжения рака37), показывают, что относительные проницаемости этих каналов для натрия, калия и кальция согласуются с более ранними наблюдениями, сделанными на целой клетке. О том, как деформация мембраны вызывает открывание каналов, пока почти ничего не известно.

370 |
Раздел III. Интегративные механизмы |
Механочувствительные волосковые клетки уха позвоночных
Наше понимание механотрансдукции продвинулось дальше всего в изучении волосковых клеток позвоночных. Механочувствительные волосковые клетки внутреннего уха реагируют на акустическую вибрацию или движения головы, вызывающие перемещение жидкости в полостях внутреннего уха. Точная форма перемещения жидкости зависит от конфигурации и состава конкретного концевого органа, задействованного в каждом процессе. В главе 18 мы обсуждаем частотно-специфичный паттерн колебаний мембран в спирали улитки и различные функции внутренних и внешних волосковых клеток для слухового восприятия. Здесь достаточно будет указать, что волосковые клетки в улитке (cochlea) стимулируются движениями жидкости в диапазоне акустической частоты — у человека от 20 до 20000 Гц. Вестибулярные концевые органы внутреннего уха построены совершенно иначе и реагируют на гораздо более низкие частоты, возникающие при движениях головы. Гравитационная нагрузка на мешочек (saccule) и маточку (utricle), создаваемая содержащей кристаллики отолитовой мембраной (otolithic membrane), обеспечивает чувствительность этих эпителиальных структур к линейному ускорению. Волосковые клетки в полукружных каналах (semicircular canals) активируются угловым ускорением при вращении головы. Независимо от типа движения, перемещение жидкости вызывает отклонение пучка модифицированных микроворсинок, или стереоцилий, которые отходят от апикальной
Рис. 17.8. Механочувствительная волосковая клетка. (А) Схема функциональной специализации волосковой клетки. (В) Электронная микрофотография волосковой клетки из внутреннего уха цыпленка.
Fig. 17.8. The Mechanosensory Hair Cell. (A) Schematic drawing highlighting the functional specializations of the hair cell. A bundle of specialized microvilli called stereocilia projects from the cuticular plate into the endolymphatic space.
In some hair cells a true cilium, the kinocilium, is found at cne side of the hair bundle. Below the nucleus, hair cells form synapses with afferent and efferent neurons. Synaptic vesicles surround a dense body opposite the ending of an afferent neuron. Efferent neurons projecting from the brainstem form cholinergic synapses. Inside the hair cell a- synaptic cistern lies in close apposition to the plasma membrane underlying the efferent contact. (B) Transmission electron micrograph of a hair cell from chick inner ear. The hair bundle was bent over during fixation. In this type of hair cell the cuticular surface is expanded. Synaptic contacts are made on the basal pole of the celL (Micrograph kindly provided by R. Michaels.)
Глава 17. Трансдукция механических и химических стимулов |
371 |
поверхности волосковой клетки. Отклонение пучка напрямую приводит к открыванию механочувствительных ионных каналов.
Структура рецепторов волосковых клеток
Волосковые клетки и окружающие их вспомогательные клетки формируют эпителиальные слои, которыми отделены друг от друга разнородные жидкие среды внутреннего уха. Базально-латеральные участки мембраны волосковых клеток омываются перилимфой, сходной по составу с обычной внеклеточной жидкостью, содержащей много натрия и мало калия (рис. 17.8). Апикальная поверхность волосковой клетки, формирующая волоски, повернута в сторону эндолимфы, раствора, сходного определенным образом с цитоплазмой и содержащего калий в высокой концентрации, а натрий и кальций — в низкой. Волосковые клетки на своих базально-латеральных поверхностях формируют синаптические контакты с афферентными волокнами. Некоторые волосковые клетки также получают синаптические входы от эфферентных нейронов ствола мозга.
На волосковых клетках разных типов, независимо от места их расположения, находится от нескольких десятков до сотен ранжированных по высоте стереоцилий (видоизмененных микроворсинок, содержащих филаменты полимеризованного актина). Самые длинные стереоцилий обнаружены на волосковых клетках полукружных каналов, самые короткие — в высокочастотной области улитки. В составе любого пучка стереоцилий расположены в порядке повышения высоты, как трубы органа или как ступеньки лестничного марша. Во многих волосковых клетках находится только один настоящий волосок, киноцилий (содержащий набор из 9 + 2 микротрубочек), который находится возле центра самого высокого ряда стереоцилий. В волосковых клетках улитки киноцилии присутствуют только в раннем онтогенезе, позже они исчезают. Стереоцилий закреплены своим основанием в покровной пластинке. Во время отклонения пучка стереоцилий ведут себя как жесткие удилища и сгибаются только в месте прикрепления38). Разнообразные боковые сочленения способствуют тому, что скопление стереоцилий перемещается как единый колосковый пучок.
Трансдукция через отклонение волоскового пучка
Уже несколько лет известно, что электрические реакции в волосковых клетках возникают благодаря деформации волоскового пучка39· 40), однако для непосредственного экспериментального подтверждения этого потребовалась разработка чувствительной аппаратуры, которая могла бы вызвать и измерить ничтожно малые перемещения при отведении от волосковых клеток. Слуховой и вестибулярный эпителий холоднокровных позвоночных, таких как черепахи и лягушки, оказался наиболее удобным для проведения таких опытов. Процедуры, использованные Кроуфордом и Феттиплейсом для стимуляции волосковых клеток в базальном сосочке (basilar papilla) черепахи (слуховой эпителий, аналог органа Корти у млекопитающих), показаны на рис. 17.9А. Базальные сосочки были извлечены из внутреннего уха и помещены в солевом растворе в камеру, смонтированную на предметном столике микроскопа. Микроэлектрод записывает потенциал мембраны волосковой клетки, в то время как стекловолокно, прикрепленное к пьезоэлектрическому манипулятору, давит на пучок волосков. Проецируя изображение стекловолокна на пару фотодиодов, можно уловить
смещения всего на 1 нм. Такой стимул производит в волосковой клетке изменение потенциала величиной в 0,2 мВ 41).
Хадспет и его коллеги детально описали процесс трансдукции в вестибулярных волосковых клетках лягушки42)--44). В одной из серий экспериментов они непосредственно продемонстрировали функциональную ориентацию волоскового пучка, варьируя направление стимуляции, осуществляемой пьезоэлектрическим манипулятором. Отклонения в сторону киноцилия деполяризовали клетку, тогда как движение от него приводили к гиперполяризации. Отклонение пучка в направлении, перпендикулярном этой оси, не вызвало изменений мембранного тока45). Результаты такого эксперимента приведены на рис. 17.9В, где величина изменения потенциала, генерируемого в волосковой клетке, изменяется в зависимости от угла отклонения пучка.
Концевые связи и воротные пружины
Какая структурная особенность волоскового пучка могла бы лежать в основе ориентаци-

372 |
Раздел III. Интегративные механизмы |
|
|
Рис. 17.9. Регистрация механотрансдукции в волосковых клетках. (А) Микроэлектрод, введенный в
изолированную волосковую клетку. (В) Рецепторные потенциалы, зарегистрированные в волосковой клетке изолированного мешочка лягушки при разных углах отклонения пучка. Положение киноцилии соответствует 0°.
Fig. 17.9. Recording Mechanotransduction in Hair Cells. (A) Microelectrodes are inserted into hair cells in an excised epithelium mounted on the stage of a compound microscope. The hair bundle is displaced by a glass fiber attached to a piezoelectric manipulator. The image of the glass fiber is enlarged and projected onto a photodiode pair so that motion causes a differential signal between them. Movements as small as 1 nm can be detected with this method. (B) Receptor potentials recorded from a hair cell in the excised saccule of a frog during bundle deflection at various angles. The kinocilium at the center of the tallest row of hairs lies at 0°. Maximal responses occur for motion toward and away from the kinocilium; no response is seen during motion at right angles to that line. (A after Crawford and
Fettiplace, 1985; В after ShotwelL Jacobs, and Hudspeth, 1981.)
онной избирательности трансдукции? Пикльз и коллеги, используя сканирующий электронный микроскоп, описали уникальный класс внеклеточных связей, соединяющих верхушки одного волоска с боковой поверхностью соседнего, более длинного волоска46). Эти концевые связи (рис. 17.10А) наблюдались только вдоль оси механической стимуляции (т. е. ориентированы вверх и вниз по «лестнице»). Положение концевых связей позволяет предположить, что они могут быть вовлечены в механотрансдукцию, а воздействие, разрывающее концевую связь, должно прекращать трансдукцию47· 48). Действительно, внеклеточные отведения указывают на то, что каналы, активируемые механическим стимулом, расположены вблизи вершины волоскового пучка49)--51).
Количественные измерения трансдукции и идентификация концевых связей были объединены в гипотезе воротной пружины (gating spring), описывающей механизм механотрансдукции в волосковых клетках. Отклонение волоскового пучка в «позитивном» направлении (в сторону более высоких волосков) увеличивает расстояние между концами волосков, растягивает воротную пружину, открывая таким образом ворота каналов трансдукции (рис. 17.10В). Когда пучок смещается в сторону, противоположную более длинным волоскам, происходит сжатие пружины, и каналы закрываются. Хотя такая схема может показаться достаточно идеализированной, однако требование прямой физической связи между механикой пучка и открыванием канала диктуется огромной скоростью, с которой в волосковых клетках происходит трансдукция, имеющая постоянную времени открывания около 40 мкс52· 53). К тому же энергетика и механика трансдукции согласуются с данной моделью. Например, можно измерить уменьшение жесткости пучка по мере того, как открываются каналы трансдукции, так,
будто это молекулярное движение действительно снимает напряжение с воротной пружины54).