

Nikolls_-_Ot_neyrona_k_mozgu
.pdfГлава 21. Функциональная архитектура зрительной коры |
473 |
|
|
Рис. 21.9. Изменение направления движения глаза при электрической стимуляции в области МТ (А) Запись движения глаза в ответ на движущийся зрительный стимул. До этого микроэлектрод был введен в область зоны МТ, и было определено предпочтительное направление движения для клеток этой области, которое отличается от направления движения данного стимула (В). При электрической стимуляции зоны МТ результирующий вектор движения отклоняется в сторону. Эти данные дают основания предполагать, что зрительные движения вычисляются как суммарный вектор нескольких предпочтительных направлений в зоне МТ.
Fig. 21.9. The Direction of Eye Movements Can Be Altered by Electrical Stimulation in Area MT. (A) Eye movements were recorded in response to a moving visual target. Earlier, an electrode had been inserted into area MT and the preferred direction of cells in that location was noted. This preferred direction differed from that of the moving target.
(B) When this location in area MT was stimulated electrically, the resulting eye movements were biased in the preferred direction of cells in the stimulated region. These results suggest that visual motion is computed as the vector sum of several preferred directions in area MT. (After Groh, Born, and Newsome, 1997.)
ентацию, совпадающую с направлением движения объекта. Но движущиеся зрительные объекты активируют не только одну подобную колонку. Чаще всего они имеют замысловатую траекторию движения, вызывая активацию большого набора нейронов, чувствительных к различным направлениями движения и к разным его углам. Стало быть, для оценки такого движения и расчета усредненного вектора движения потребуются нейрональные вычисления. Нейрональные вычисления, осуществляемые колонками, чувствительными к направлению движения объекта, изучались Ньюсомом с коллегами, с использованием электрической микростимуляции для управления движения глазом у тренированных обезьян44). При помощи микроэлектрода регистрировалось предпочтительное направление движения для данной колонки клеток, а затем этот электрод использовался для инъекции небольшого по величине тока для активирования данной колонки в то время, когда осуществлялось следящее движение глазом (рис. 21.9). После этого проводили сравнение следящего движения глаза без стиму-
ляции и при стимуляции МТ, для того чтобы узнать, каким образом происходит сложение двух различных направлений движения в пределах зрительного поля. В контроле глаз строго следует за объектом (рис. 21.9А). При стимуляции электрическим током колонки МТ, предпочтения относительно движения которой отличались от направления движения объекта, результирующее движение глаза было средним между двумя этими направлениями (рис. 21.9В). Вывод: траектория движения используется для расчета среднего вектора для активированных колонок, отслеживающих направление движения. Привлекательной особенностью таких экспериментов является то, что поведение обезьяны (движение глаз) представляет собой тип анализа, выполняемый в высших корковых центрах Уже давно предполагалось, что подобный расчет среднего вектора используется при выполнении резкого отрывистого движения глаз45).
Цветовое зрение
Мелкоклеточные нейроны зрительного тракта несут информацию о тонких деталях формы

474 |
Раздел III. Интегративные механизмы |
Рис. 21.10. (См. цветную вклейку после с. 640.) Организация чувствительного к цвету рецептивного поля клеток мелкоклеточных (parvocellular) слоев в ЛКТ обезьяны. (А) Организация рецептивного поля с красным "on"-центром и зеленым "off"-фоном. Подобного типа клетка наилучшим образом отвечает на маленькое или большое красное пятно на нейтральном фоне. Красное пятно на зеленом фоне, однако, вызывает относительно слабый эффект. (В) Пример организации рецептивного поля с различной спектральной чувствительностью
Fig. 21.10. Receptive Field Organization of Cells in parvocellular layers of the monkey LGN, responding to color. (A) Receptive field organization with a red "on" center and a green "off' surround. A cell of this sort would fire best in response to a small or large red spot on a neutral background. A red spot on a green background would be relatively ineffective. (B) Example of receptive field organizations with various spectral sensitivities. (After Wiesel and Hubel 1966; Hubel 1988.)
и цвета объектов. На уровне колбочек мы видели четкую корреляцию между нейронными сигналами и длиной волны падающего на сетчатку света (глава 19). Колбочки красного, зеленого и синего типа предпочтительно поглощают свет в области длинно-, средне--и коротковолнового спектра. В принципе, при помощи сравнения активности каждого типа колбочек, нервная система может рассчитать длину волны света. Однако таким ли образом осуществляется восприятие цвета центральным зрительным анализатором?
Конвергенция сигналов от колбочек начинается с уровня горизонтальных клеток, преобразующих эти сигналы в цветовой код, и продолжается в дальнейшем в ганглиозных клетках и мелкоклеточной части латерального коленчатого тела. Свойства подобных ганглиозных клеток и клеток ЛКТ, использующих цветовой код, показаны на рис. 21.10. Значительной трансформации свойств рецептивного поля при прохождении сигнала от зрительного нерва до клеток ЛКТ не наблюдалось46). Рецептивные поля клеток ЛКТ на рис. 21.1 ОА имеют концентрическую форму, с красным "on"-центром и зеленой "off"-периферией. Маленькое красное пятно, освещающее центр, вызывает бурный разряд; большее по размеру зеленое пятно, нанесенное не в центральной области, приводит к торможению. Подобного рода клетка отвечает наилучшим образом на красный цвет на нейтральном или голубом, но не на зеленом фоне. Она имеет традиционные свойства «центр-периферия» при ответе на вспышки белого цвета. Другие клетки имеют желто-голубые антагонистические зоны (желтый как смесь красного и зеленого цвета). Разные типы организации рецептивного поля «центрпериферия», наблюдаемые в мелкоклеточных слоях ядра латерального коленчатого тела обезьян, показаны на рис. 21.10В.
Красно-зеленые и желто-голубые нейроны представляют собой примеры клеток, воспринимающих противоположные цвета. Они анализируют длину волны, сравнивая сигналы, поступающие на них от колбочек именно таким образом, как представляли себе Янг и Гельмгольц (глава 19). Красный, зеленый, голубой, желтый, черный и белый шар на бильярдном столе вызывает в этих клетках определенные, уникальные сигналы, которые затем передаются в мозг.
Пути цветного зрения
Картина последовательных шагов коркового анализа цвета и его восприятия была составлена на основании экспериментов Зеки, Хьюбеля, Доу, Ланда и их коллег. Как уже упоминалось, пути передачи информации о цвете отделены, в основном, от путей, занимающихся анализом других свойств, таких как пространственная глубина изображения, движение, контраст и форма. Зеки показал, что мелкоклеточный путь, идущий от зоны V1 через V2 к зоне V4, имеет в своем составе большое количество клеток, кодирующих цветовую информацию47). Доказательство ключевой роли зоны V4 в цветовом зрении было получено при помощи позитронно-эмиссионной томографии и функционального магнитного резонанса при изучении здоровых людей. При проецировании цветовых паттернов на сетчатку повышенная активность была отмечена в области, совпадающей с зоной V448)-50) Отдельное восприятие цвета и формы изображения наиболее убедительно демонстрируется в тех редких случаях, когда пациенты страдают потерей цветового зрения из-за изолированного повреждения головного мозга (церебральная ахроматопсия)51·52). Например, описан случай, когда пациент перенес двустороннюю травму мозга в той части, которая располагается перед зоной V1, что примерно соответствует области V453). До повреждения у него было нормальное цветовое зрение, но после травмы он потерял способность различать цвета. Он знал из своего предыдущего опыта, что земляника имеет
Глава 21. Функциональная архитектура зрительной коры |
475 |
красный цвет, а банан — желтый, но после повреждения все виделось ему как будто в чернобелом кино. Другие функции, такие как память и распознавание форм, были лишь незначительно повреждены, и он был способен продолжать свою работу таможенного инспектора(!). При демонстрации ему какого-либо объекта он мог описать, какой цвет должен иметь этот объект, но не мог сопоставить этот цвет с предложенной цветовой шкалой. Однако никаких дефектов ни в речи, ни в распознавании объектов не наблюдалось, только лишь в самом восприятии цветов.
Психофизические исследования у нормальных людей подтверждают отделение информации о цвете от другой информации сразу после восприятия. Подробные описания этого даны в статьях55) и обзорах Зеки40), Хьюбеля56) и Ливингстона57). Например,трудно, или даже невозможно, выделить структуру или форму изображения, если не происходит активация крупноклеточных путей зрительного анализатора, воспринимающих зоны контраста на изображении, обычно представленные различными степенями яркости и тени. Мелкоклеточная система, с ее акцентом на восприятие цвета и высокое пространственное разрешение, имеет ограниченные возможности по восприятию формы предметов. Следовательно, цветное изображение сложной структуры с многими компонентами, отражающими зоны с одинаковом уровнем освещенности, для мелкоклеточной системы представляется таким, что не содержит конкретных форм. Это происходит потому, что не работает крупноклеточный путь. Подобным же образом наши ощущения глубины изображения и движения также могут терять свою эффективность, если контраст черно-- белого изображения недостаточен для активации крупноклеточных путей. Впечатляющей демонстрацией этого является перемещение рисунка из зеленых и красных полосок вдоль телевизионного экрана. Интенсивность каждого цвета может быть подобрана так, что полоски становятся эквилюминентными (т. е. каждая красная или зеленая полоска излучает такое же эффективное количество света, как и соседняя, хотя и на другой длине волны). Мы попрежнему видим цветовые полоски, но кажется, что они перестали двигаться.
Цветовое постоянство
Основная проблема в нашем понимании цветного зрения — это понять, каким образом кора определяет, какого цвета тот или иной объект зрительной сцены. В нашем мозге подобного рода расчеты так удачно запрограммированы, что мы интуитивно не осознаем, что здесь может быть какая-либо проблема. Разумеется, иллюстрации голубого цвета в этой книге выглядят голубыми потому, что они отражают свет на короткой длине волны. Из всего, сказанного до сих пор, можно представить себе, что цвета, которые мы видим, определяются просто и непосредственно длиной волны света. Однако для Гельмгольца это не было так очевидно58). Он указывал, что яблоко, которое мы видим днем, на закате и в свете свечи выглядит красным. Однако свет, отраженный от его поверхности, содержит гораздо больше красного цвета на закате и гораздо больше желтого в свете свечи. Каким-то образом мозг «приписывает» красный цвет яблоку и не меняет своего восприятия даже при очень различных условиях. Изображение яблока в мозге как бы «не принимает в расчет освещение».
Сходным примером является тон двух корректно экспонированных фотографий, сделанных на одной и той же пленке при дневном свете и в комнате с искусственным светом от электрических ламп. Цвета в дневном свете выглядят более реалистичными, а на фото, сделанном в помещении, имеют больше желтого цвета. Однако мы, тем не менее, совсем не осознаем этой желтизны, когда искусственно освещаем комнату. (Этот феномен до недавнего времени наблюдался очень часто; сейчас вспышки, присутствующие почти на каждом фотоаппарате, имеют спектр, близкий к спектру дневного света). Биологические преимущества цветового постоянства очевидны: зеленые ягоды не должны превращаться в красные на закате; розовые губы не должны становиться желтыми в свете свечи.
Впечатляющая демонстрация цветового постоянства была разработана Ландом59· 60), что послужило мощным стимулом для нейробиологических исследований в области цветового зрения. Его демонстрация показала, что то, каким мы видим цвет объекта, существенно зависит от света, отраженного от всего изображения, а не только от самого объекта. Мы не можем определить цвет — желтый, зеленый, голубой или белый — для какой либо области, только определяя длину волны отраженного от этой области света. Нам также необходимо знать композицию света, отраженного от соседних областей. Такой стран-

476 |
Раздел III. Интегративные механизмы |
Рис. 21.11. (См. цветную вклейку после с. 640.) Рецептивное поле клетки двойного противопоставления, свойства которой помогают объяснить постоянство цвета. Клетка подобного типа отвечает наилучшим образом на маленькое красное пятно (R), проецируемое на зеленом фоне (G) (верхняя запись), или на зеленое кольцевидное освещение (G) (нижняя запись).
Fig. 21.11. Receptive Field of a Double-Opponent Cell the properties of which could help to explain color constancy. A cell of this type responds best to a small red spot shining on a green background (upper record), or to green annular illumination (lower record). (After Daw, 1984.)
ный вывод, известный как феномен Ланда, кажется противоположным тому, что нам говорит наша интуиция. Так же как для черного и белого, мозг формирует восприятие цвета, сравнивая свет, падающий на различные области сетчатки, вместо того, чтобы измерять абсолютную яркость и длину волны в одном ее месте. Скорее всего, это как если бы в коре проводилось тотальное сравнение контраста на всех границах изображения для трех различных изображений, видимых через коротко-, средне- и длинноволновые фильтры.
Невозможно дать всеобъемлющее и удовлетворительное описание феномена Ланда в терминах свойств рецептивных полей клеток, кодирующих цвета в областях V1, V2 и V4. Однако один тип клеток, известный как «клетки двойного противопоставления» (double opponent cells), имеет свойства, которые могут принимать участие в восприятии цветового постоянства. Первоначально они были описаны Доу62· 63) в сетчатке золотой рыбки. Затем подобные клетки были обнаружены в коре приматов, но не в ядрах латерального коленчатого тела или в сетчатке28· 64)--66). Следовательно, они участвуют в более поздних стадиях переработки информации о цвете. Вкратце, такие клетки имеют рецептивные поля примерно концентрической формы в виде «центр-периферия», имеющие красно-зеленый или желтоголубой антагонизм (рис. 21.11). Но, в отличие от клеток цветного противопоставления в ЛКТ,
вклетках двойного противопоставления каждый цвет вызывает антагонистичные эффекты, как
вцентре, так и в области периферии. Следовательно, при освещении красным цветом в центре рецептивного поля происходит "on"-разряд, красное же освещение периферии приводит к "off"-разряду. Зеленый цвет в области периферии приводит к "on"-разряду, а в центре — к "off".
Предположим, что мы вызываем разряды при помощи маленького красного пятна в центре рецептивного поля такой клетки, используя в качестве фона монотонное белое освещение. Если мы сейчас увеличим долю красного цвета в монотонном освещении, то уровень сигнала изменится только незначительно: увеличенное возбуждение центральной зоны красным цветом будет компенсироваться более сильным торможением в ответ на освещении периферических зон красным. В самом деле, баланс сигналов с колбочек, воспринимающих красный, зеленый и голубой цвета, различен для центра и периферии, а также колеблется от клетки к клетке в области первичной зрительной коры (V,), таким образом можно наблюдать непрерывный переход антагонистических оттенков67·68). Это противоречит цветовым предпочтениям нейронов ЛКТ, которые в значительной степени совпадают с основными воспринимаемыми цветами. Можно предположить, что длинные горизонтальные связи между пятнами (см следующий раздел) играют определенную роль в пространственных взаимосвязях, которые позволяют объяснить феномен Ланда.
§3. Интеграция зрительной информации
Горизонтальные связи в пределах первичной зрительной коры
Схема обработки зрительной информации, подобная той, что показана на рис. 21.7, представляет собой рабочую модель, которая помогает придать нашим представлениями более организованный вид. Однако разделение крупноклеточного и мелкоклеточного пути — для определения контраста, движения и глубины изображения, с одной стороны, и цвета и фона — с другой — ни в коей мере не является полным. Взаимодействие между ними обнаруживается даже в области V,, где сигналы от крупноклеточных клеток можно обнаружить в зоне пятен и между пятнами20, 30, 69, 70). Более того, только зоны V1 и V2 четко определены и относительно их границ имеется согласие; дополнительные же зоны ассоциативной зрительной коры не имеют четко очерченных границ. Свойства рецептивных полей клеток,
Глава 21. Функциональная архитектура зрительной коры |
477 |
расположенных в этих зонах, могут сильно варьировать и различные типы зрительных полей могут быть представлены не в столь четко организованном порядке.
В самой зоне V1 было описано большое разнообразие связей, что предполагает наличие более сложных принципов организации, чем ранее предполагалось. Использование классических методов окраски, таких как окраска (импрегнация) по Гольджи, выявляет доминирование нейронных отростков, которые направляются, в основном, перпендикулярно поверхности коры из слоя в слой. При помощи внутриклеточных инъекций красителей было показано, что кортикальные нейроны имеют также длинные горизонтальные отростки, которые простираются латерально от колонки к колонке (рис. 21.12А)71)--78). Соединения, подобные этим, дают большой вклад в синтез удлиненных рецептивных полей простых клеток слоя 6 зоны V1 : рецептивные поля клеток слоя 5 комбинируются и добавляются конец в конец к полям простых клеток слоя 6 при помощи длинных горизонтальных аксонов. Было обнаружено большое количество простых и комплексных клеток с длинными горизонтальными отростками, имеющими длину более 8 мм, образующих сверхколонки. Отдельный нейрон, таким образом, может интегрировать информацию с целой зоны поверхности сетчатки в несколько раз превосходящей размеры рецептивного поля, измеряемого стандартными методами79).
Особенный интерес представляет то, что соединения образуются между колонками, которые имеют сходные ориентационные особенности. Доказательства таких особых соединений были получены при помощи двух дополнительных методов. Во-первых, когда метки были введены в
одну колонку, они транспортировались в удаленную сверхколонку, имеющую те же ориентационные предпочтения (рис.21.12В). Во-вторых, при помощи перекрестной корреляции паттернов активности нейронов, имеющих одни и те же ориентационные предпочтения, но расположенных в разных удаленных друг от друга колонках, можно сделать вывод, что между ними имеются функциональные связи30, 80, 81). Более того, после повреждения сетчатки, кортикальные клетки, лишенные сигнала, также демонстрируют ответы на
удаленные стимулы, которые располагаются вне пределов их «нормальных» рецептивных полей28).
Рецептивные поля обоих глаз, конвергирующие на кортикальных нейронах
Когда мы смотрим на объект одним или двумя глазами, мы видим только одно изображение, даже если размер и расположение проекции объекта немного отличается на двух сетчатках. Интересно, что еще более 100 лет назад Иоханес Мюллер предположил, что отдельные нервные волокна от обоих глаз могут пересекаться и образовывать связи с одними и теми же клетками в ЦНС. Таким образом, он почти предвидел результаты, полученные Хьюбелем и Визелем82· 83). Они обнаружили, что около 80 % всех кортикальных нейронов в зрительных областях мозга кошки получают сигналы от обоих глаз. Поскольку нейроны, располагающиеся
вразличных слоях ЛКТ, преимущественно иннервированы либо одним, либо другим глазом, формирование перекрестного взаимодействия между различными глазами становится возможным только в коре. Как уже упоминалось ранее, разделение происходит в слое 4 первичной зрительной коры, где каждая простая клетка получает сигнал только от одного глаза, игнорируя другой. Смешивание сигнала от двух глаз происходит на следующих этапах переключения, то есть в слоях, расположенных глубже (по направлению к белому веществу) и
вслоях, более близких к поверхности коры.
Исследование рецептивных полей клеток, получающих бинокулярную информацию, показывает, что (1) рецептивные поля их обычно находятся в абсолютно эквивалентных частях зрительного поля обоих глаз, (2) они имеют одинаковую предпочтительную ориентацию и (3) соответствующие зоны рецептивных полей дополняют эффекты друг друга. Синергичное действие двух глаз на примере простой клетки показано на рис. 21.13. Освещение "of"-зоны левого глаза суммируется с освещением "of"-зоны правого глаза. Одновременное освещение в антагонистических зонах обоих глаз уменьшает текущую активность и усиливает "off"- разряды. Подобные клетки отвечают сигналами на одинаковые изображения в обоих глазах.
Для восприятия глубины изображения существует иная бинокулярная специализация рецептивных полей84). Объект, находящийся за плоскостью фокуса, проецируется в неодинаковые зоны двух сетчаток. Нейроны, обладающие свойствами воспринимать глубину
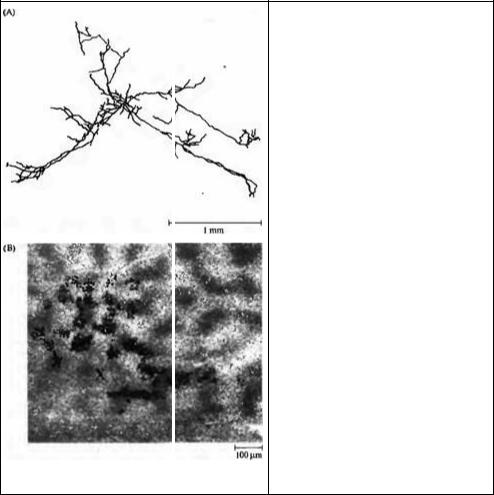
478 |
Раздел III. Интегративные механизмы |
Рис. 21.12. Горизонтальные связи в зрительной коре. (А) Вид поверхности пирамидальной клетки зоны V1 кошки после введения пероксидазы хрена. Отростки простираются примерно на 3 мм вдоль поверхности коры. Тонкие веточки и синаптические бутоны данного нейрона обнаруживаются в нескольких отдельных кластерах, отдаленные друг от друга на расстояние 800 мм и более. (В) Микросферы с метками были введены в область, где клетки имеют предпочтение к вертикальной ориентации (помечено черным "X"). Микросферы захватываются терминалями аксона и транспортируются ретроградно в тела клеток, проецирующих свои отростки в область введения. Колонки вертикальной ориентации были также помечены, используя деоксиглюкозу, во время стимуляции глаза вертикально ориентированными полосками света. Микросферы с метками были обнаружены в зонах, помеченных деокси глюкозой, что говорит о наличии горизонтальных связей между клетками одной и той же ориентационной чувствительности.
Fig. 21.12. Horizontal Connections in Visual Cortex.
(A) Surface view of pyramidal cell in cat V,, after labeling with horseradish peroxidase. The processes extend for nearly 3 mm across the cortical surface.
Fine branches and synaptic boutons of this neuron occurred in several discrete clusters separated by 800 mm or more. (B) Labeled microspheres were injected into a region where cells had vertical orientation preference (large black "X"). The microspheres are taken up by axon terminals and transported back to the somata of cells projecting to the injection site. Vertical orientation columns were also labeled using deoxyglucose during stimulation with vertically oriented bars of light. The labeled microspheres are found in deoxyglucose-labeled areas, showing that horizontal connections occur between cells with the same orientation specificity.
(A from Gilbert and Wiesel, 1983; В from Gilbert and Wiesel, 1989; kindly provided by C. Gilbert.)
трехмерного изображения, были обнаружены в первичной и ассоциативной зрительной коре. Для таких клеток оптимальным стимулом является определенным образом ориентированная полоска, расположенная впереди от плоскости фокуса (для одних клеток) или позади ее (для других)64· 85· 86). При представлении этой полоски только одному глазу или обоим глазам, однако в пределах плоскости фокуса, сигналы не вызываются. Для того, чтобы клетка ответила разрядами, необходимо, чтобы изображение было различным на обеих сетчатках. Однако такое различное изображение на сетчатках может приводить к активации комплексных клеток первичной зрительной коры и к отклонению глаза для того, чтобы сфокусироваться на объекте87). Восприятие глубины осуществляется в высших корковых зонах. Например, кластеры нейронов, имеющие предпочтения для подобного рода различных бинокулярных изображений, были обнаружены в ассоциативной зрительной коре V5 (зона МТ)88). При электрической стимуляции этих нейронов у тренированных обезьян нарушалось восприятие глубины изображения.
Связи, объединяющие правое и левое зрительные поля
Отдельная проблема касается того, каким образом две коры (левая и правая) связаны друг с другом и как они работают совместно для формирования единого изображения тела

Глава 21. Функциональная архитектура зрительной коры |
479 |
Рис. 21.13. Бинокулярная активация простого кортикального нейрона, имеющего идентичные рецептивные поля в обоих глазах. Одновременное освещение корреспондирующих "on" зон (+) правого и левого рецептивного поля более эффективно, чем освещение только одного из них (верхние три записи). Аналогичным же образом, стимуляция "off"-зон (- ) обоих глаз усиливает "off"-разряды друг друга (нижние три записи). Напротив, клетки, которые занимаются восприятием глубины изображения, имеют рецептивные поля, расположенные в различных зонах зрительного поля для разных глаз. Подобные клетки требуют, чтобы полоска света находилась дальше или ближе к глазу, чем плоскость фокуса
Fig. 21.13. Binocular Activation of a simple cortical neuron that has identical receptive fields in both eyes. Simultaneous illumination of corresponding "on" areas (+) of right and left receptive fields is more effective than stimulation of one alone (upper three records). In the same way, stimulation of "off" areas (-) in the two eyes reinforces each other's "off" discharges (lower three records). In contrast cells used tor depth perception have receptive fields in the two eyes in disparate regions of the visual field. Such cells require that the bar is farther from or closer to the eye than the plane of focus. (After
Hubel and Wiesel 1959.)
и окружающего мира. Каждое полушарие воспринимает только одну половину окружающего нас мира. Это в равной степени справедливо также для восприятия прикосновения, положения тела и является основной особенностью нашего восприятия. Естественным является интерес к тому, что же происходит на их границе. Каким образом две стороны нашего мозга смешивают вместе мир, расположенный справа и слева, таким образом, что мы не можем заметить даже какого-либо намека на «шов» или прерывистость восприятия?
Самым очевидным способом сохранения постоянства восприятия является объединение правого и левого зрительных полей вместе на границе. Для того, чтобы достичь этого, клетка в правом полушарии, которая отвечает на горизонтальную полоску в центре поля зрения, должна каким-то образом быть связана с подобной же клеткой в левом полушарии, которая отвечает за продолжение этой самой полоски. Подобные взаимодействия позволили бы сформировать полную картину окружающего пространства с минимальным количеством связей между двумя полушариями. С другой стороны, бесцельно было бы связывать рецептивные поля, расположенные на границах, направленных в противоположные стороны. Высоко специализированные связи между нейронами с рецептивными полями, расположенными строго по средней границе, были обнаружены в экспериментах. Связи между этими полями идут из одного полушария в другое через мозолистое тело89).
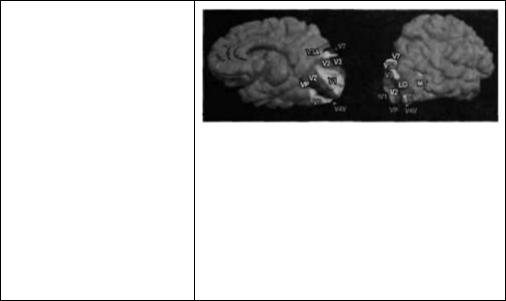
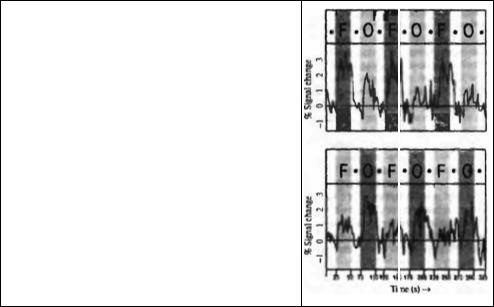
Глава 21. Функциональная архитектура зрительной коры |
481 |
Рис. 21.15. (Рис. 21.15А и 21.15В (срезы зон коры) см. на цветной вклейке после с. 640.) Зоны коры, ответственные за распознавание лиц, выявленные при функциональном магнитно резонансном исследовании. При этом исследовании пациенту демонстрировались либо изображения лиц, либо другие изображения (например, ложка). При демонстрировании субъекту большего количества изображений лиц (но не других предметов) было зарегистрировано значительное и длительное повышение активности в зоне, расположенной в области веретенообразной борозды (слева на верхней сканограмме). Объекты, не являющиеся лицами, вызывали двустороннюю стимуляцию в областях, расположенных сзади (показано снизу). Усредненные процентные значения изменения сигнала в показанных областях даны справа от сканограмм. Вертикальные темные полоски показывают момент демонстрации пиц (F), или других изображений (0).
Fig. 21.15. Face-Specific Regions of Cortex, as revealed by functional MRI. During fMRI scanning, a subject was shown either faces or nonface objects (e. g., a spoon). A region in the right fusiform gyms (left side of fMRI) was identified in which pictures of faces, more than objects, produced significant and consistent increases in activity (top). Nonface objects produced bilateral stimulation of further posterior regions (bottom). The average signal percent change in the outlined areas is shown to the right of each fMRI. Vertical shading indicates the presentation of faces (F) or nonface objects (0). (From Kanwisher,
McDermott, and Chun, 1997.)
клетки, на которых конвергируют все большие и большие объемы информации об объектах, появляющихся в поле зрения. В самом деле, в зрительных областях более высокого порядка при помощи микроэлектродной регистрации были обнаружены нейроны, которые отвечают специфическим образом на лица102). При регистрации работы клеток было подтверждено, что определенный локус в области затылочно-височной коры (фузиформная извилина) активируется избирательно при просмотре изображений лиц, а не других объектов103). Как можно видеть на верхней «сканограмме» (фронтальный срез) на рис. 21.15, зона коры, отмеченная зеленым цветом, активировалась при просмотре изображений лиц, в то время как другие объекты, не являющиеся лицами (например, ложка), активировали билатеральные зоны, расположенные более каудально. Распознавание лиц может затрагивать и другие области, например центр языка. У правшей правая фузиформная извилина активировалась предпочтительно или исключительно при предъявлении изображений лии. У двух левшей, при аналогичном тесте, происходила активация этой извилины с левой стороны.
Определенная локализация области распознавания изображений человеческих лиц также подтверждается клиническими данными, когда возникает нарушение только этой, и никакой другой функции обработки зрительной информации. Такое нарушение называется прозопагнозия (prosopagnosya)109· 110). В одном таком случае человек, имеющий высокие интеллектуальные способности и хорошую память, не был способен распознавать лица, причем даже лицо своей собственной жены111). Он рассказывал: «Как-то в клубе я увидел какого-то странного субъекта, который удивленно таращился на меня. Я спросил официанта, кто это? Вы будете смеяться. Это я смотрел на себя в зеркало».
Потеря способности к распознаванию может распространяться и на другие категории, когда, например, человек, наблюдающий за птицами, утрачивает способность различать отдельные виды птиц, а делающий ставки на лошадей игрок во время забега не способен отличить одну лошадь от другой. Неврологические и невропатологические исследования показали, что прозопагнозия связана с повреждениями справа и, иногда, с двух сторон затылочно-височной коры112). Другие виды зрительных стимулов также способны вызывать определенные паттерны активности в затылочно-височной коре. Например, последовательности печатных букв (в виде строк) вызывают предпочтительную активацию в нижнезатылочной борозде левого полушария113). Соответственно, повреждения в области затылочно-теменной коры приводят к полной
482 |
Раздел III. Интегративные механизмы |
неспособности воспринимать печатный текст («чистая» алексия)114). Являются ли специализированные зоны зрительной коры врожденными, или они появляются с опытом? Хотя можно себе представить, что «нейроны для лиц» могут закладываться в онтогенезе, такое вряд ли возможно для печатного текста. Скорее всего, кора самоподстраивается под важные стимулы на протяжении всей жизни организма. В самом деле, зоны распознавания лиц в коре также активируются, когда эксперту по наблюдению за птицами показывают картинки птиц115). Формируются ли специфические регионы коры в результате долгой практики? Разрешение и воспроизводимость результатов в методах, позволяющих регистрировать активность клеток, дает нам основания полагать, что уже скоро мы сможем сами непосредственно наблюдать подобного рода изменения, подобно тому, как мы наблюдаем их в двигательной коре во время тренировок (глава 22). Развитие и пластичность неокортекса обсуждается далее, в главе 25.
Выводы
·Нейроны первичной зрительной коры организованы на основе предпочтения сигналов от одного определенного глаза (глазное доминирование) и ориентационной избирательности.
·Расположение колонок глазного доминирования и ориентационных «волчков» может быть обнаружено при помощи регистрации активности нервных клеток оптическими методами с поверхности мозга. Изоориентационные контуры стремятся пересекать зоны глазного доминирования под определенными углами, и каждая зона ориентирования располагается между двумя колонками глазного доминирования.
·Крупноклеточные, мелкоклеточные и кониоклеточные пути образуют параллельные каналы, несущие информацию от сетчатки в зрительную кору. Крупноклеточные нейроны чувствительны к движению
и контрасту. Мелкоклеточные нейроны сигнализируют о пространственных деталях изображения и его цвете. Кониоклеточные нейроны переносят цветовую информацию непосредственно к участкам коры, выявляемым как «пятна» активности цитохромоксидазы.
·«Пятна» цитохромоксидазы располагаются в центре каждой глазодоминантной колонки и представляют собой области синтеза сигналов в первичной зрительной коре (V1).
·Чередующиеся полоски коры с высокой и низкой активностью цитохромоксидазы в области V2 особым образом взаимосвязаны с подобными же полосками в области V1.
·Распознавание движения обеспечивается нейронами V5 (зона МТ) париетальной коры.
·Зона V4 в височно-затылочной области содержит в основном нейроны, кодирующие цветовую информацию.
·Клетки двойного противопоставления (double-opponent cells) в зрительной коре имеют свойства, играющие важную роль в восприятии феномена постоянства цвета.
·Интеграция рецептивных полей в коре обеспечивается длинными горизонтальными аксонами, которые соединяют между собой колонки клеток, имеющих близкие свойства.
·Большинство нейронов коры получает сигналы от соответствующих точек зрительного поля обоих глаз, но некоторые нейроны отвечают на стимулы, расположенные в различных точках двух сетчаток. При помощи подобных отличий в восприятии изображения двумя глазами в области МТ происходит стереоскопическое восприятие глубины изображения.
·Функциональные магнитно-резонансные исследования позволяют провести картирование зон активности в пределах первичной и вторичной зрительной коры, а также в более высокоспециализированных областях коры человека.