

Nikolls_-_Ot_neyrona_k_mozgu
.pdfГлава 15. Клеточные механизмы интеграции и поведения |
323 |
го» воздействия хорошо объясняет тот факт, что при давлении на кожу пиявки сначала возникает сокращение, а потом выпрямление сегментов.
Сокращение стенок тела возникает быстро и длится недолго, в то время как распрямление (erection) мышечного кольца происходит более медленно и длится дольше. Такая реакция хорошо объясняется синаптическими потенциалами, показанными на рис. 15.6, которые вызывают облегчение на определенных частотах и управляют последовательной активацией двух мотонейронов. Когда N клетка посылает серию импульсов в ответ на физиологический раздражитель, синаптические потенциалы в химических синапсах могут быть обнаружены как в L, так и в АЕ мотонейронах. Эти синаптические потенциалы проходят фазы облегчения и депрессии. Облегчение (facilitation), однако, значительно более выражено и дольше длится в синапсах АЕ мотонейронов. Большое количество косвенных экспериментов позволяют предполагать, что облегчение и депрессия зависят от количества медиатора, высвобожденного из терминалей N клеток. Сходные эффекты были обнаружены в клетках-мишенях, иннервируемых кортикальными нейронами46).
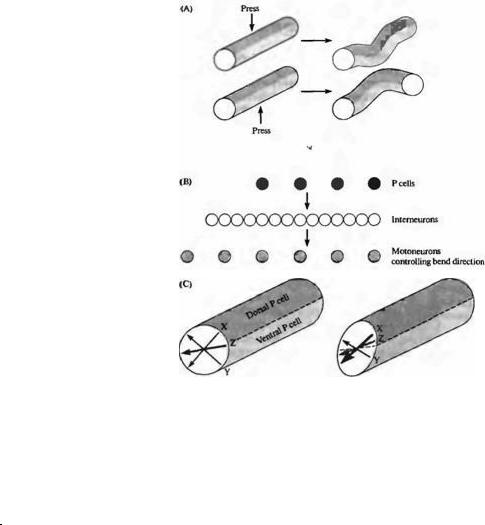
У пиявок и Aplysia47, 48) имеются и более медленные пути проведения сигнала, посредством интернейронов, которые идут параллельно основному пути. Они служат для координации более сложных движений в определенную сторону в ответ на механическое раздражение. Например, Кристан и его коллеги показали, что в случае давления на одну сторону пиявки возникает сгибание тела. Направление сгибания зависит от того, на какой участок тела наносится раздражение49· 50). Рефлекс сгибания способствует развороту тела от раздражителя. Относительная частота активности четырех Ρ нейронов, которые распознают место нанесения раздражения, анализируется сетью из 25-30 интернейронов, контролирующих сгибание тела в нужном направлении (рис. 15.7). Это достигается путем сокращения одних мышц и расслабления других. То, каким образом происходит активация двигательных нейронов, зависит от результатов работы этой нейронной сети. Интересно, что в стоматогастрическом ганглии омаров51)--53) любой интернейрон может быть вовлечен в анализ большого количества движений.
Мембранный потенциал, пресинаптическое ингибирование и освобождение медиатора
У пиявки насосную функцию кровеносных сосудов, составляющих сегментарные «сердца», контролируют мотонейроны ЦНС. Циркуляция крови зависит от перистальтических волн, распространяющихся вдоль тела животного, и скоординированного насосного механизма по обеим сторонам тела. В хорошо изученных нейронах пиявки, управляющих частотой сокращения «сердец», ритмическая активность формируется путем циклического изменения мембранного потенциала за счет синапсов вставочных нейронов24). Это, в свою очередь, регулирует процесс освобождения медиатора в двигательных клетках. Деполяризация и гиперполяризация на пресинаптическом уровне уже в течение короткого времени вызывают значительные реакции54). В частности, длительная деполяризация пресинаптической терминали в синапсе между интернейроном и двигательным мотонейроном «сердца» приводит к тому, что в ответ на потенциал действия выделяется больше медиатора. И наоборот, если пресинаптическая терминаль гиперполяризуется, высвобождается меньшее количество передатчика.
Сдвиг потенциала покоя даже на 5 мВ (от —40 до —35 мВ) может вызвать трехкратное увеличение количества высвобождаемого медиатора, в то время как амплитуда и продолжительность потенциала действия не так явно меняются в ответ на подобное изменение МП. Нервная терминаль периодически гиперполяризуется в результате тормозного влияния отдельных интернейронов, что значительно уменьшает количество квантов медиатора, выделяющихся в ответ на приходящий импульс. Это является примером того, как осуществляется пресинаптическое ингибирование при гиперполяризации, механизм которой можно объяснить путем анализа квантового состава. Торможение так реализовано во времени, чтобы обеспечивать расслабление сосудов тела с одной стороны во время сокращения их на контралатеральной стороне. Аналогичное влияние величины мембранного потенциала на пресинапсе на секрецию медиатора можно более тщательно изучить на синапсах, образующихся между нейронами в культуре клеток55).
324 |
|
|
|
|
|
|
|
|
Раздел III. Интегративные механизмы |
|
|
|||
Рис. |
15.7. |
Интеграция |
|
информации |
|
|
|
|||||||
интернейронами |
при |
осуществлении |
|
|
|
|||||||||
рефлекса сгибания тела у пиявки. При |
|
|
|
|||||||||||
давлении на какую-либо часть тела |
|
|
|
|||||||||||
пиявки |
|
возникает |
|
сгибательный |
|
|
|
|||||||
рефлекс, в результате которого эта |
|
|
|
|||||||||||
часть тела перемещается в сторону от |
|
|
|
|||||||||||
действия |
раздражителя. |
Сенсорные |
|
|
|
|||||||||
нейроны прикосновения (Т) и давления |
|
|
|
|||||||||||
(Р) посылают сигналы к серии |
|
|
|
|||||||||||
интернейронов, которые, в свою |
|
|
|
|||||||||||
очередь, образуют возбуждающие и |
|
|
|
|||||||||||
тормозные связи с мотонейронами. (А) |
|
|
|
|||||||||||
При |
|
активации |
|
Ρ |
|
клетки, |
|
|
|
|||||
иннервирующей |
дорзальную |
область |
|
|
|
|||||||||
кожи, |
|
|
продольные |
|
|
мышцы |
|
|
|
|||||
непосредственно |
под |
|
местом |
|
|
|
||||||||
прикосновения сокращаются, а на |
|
|
|
|||||||||||
противоположном |
|
|
|
конце |
|
|
|
|||||||
расслабляются |
|
за |
|
счет |
|
|
|
|||||||
торможения. |
Аналогичным |
образом |
|
|
|
|||||||||
при |
|
давлении |
на |
|
вентральную |
|
|
|
||||||
поверхность |
|
тела |
|
сокращаются |
|
|
|
|||||||
продольные |
|
мышцы |
|
брюшной |
|
|
|
|||||||
стороны, а мышцы дорзальной |
|
|
|
|||||||||||
стороны расслабляются. (В) Схема |
|
|
|
|||||||||||
соединений между 4-мя Ρ клетками и |
|
|
|
|||||||||||
интернейронами, которые, в свою |
|
|
|
|||||||||||
очередь, активируют и ингибируют |
|
|
|
|||||||||||
двигательные нейроны, участвующие в |
|
|
|
|||||||||||
сгибании тела. (С) При активации |
|
|
|
|||||||||||
обеих Ρ клеток одновременно (в |
|
|
|
|||||||||||
случае |
одновременного |
давления |
в |
|
|
|
||||||||
точках |
X и |
Y или при |
давлении |
в |
|
|
|
|||||||
точке |
|
Z) интернейроны |
интегрируют |
|
|
|
||||||||
информацию |
|
и |
|
рассчитывают |
|
|
|
|
||||||
|
|
These in turn make excitatory and inhibitory connections on motoneurons. |
||||||||||||
правильный |
|
вектор |
|
сгибания. |
|
|||||||||
Направление |
сгибания |
может быть |
|
(A) When the Ρ cell innervating dorsal skin is |
activated by |
pressure |
||||||||
рассчитано |
|
на |
основе |
разностей |
(arrow), the longitudinal muscles immediately |
underneath |
the skin |
|||||||
частот импульсации обеих Ρ клеток. |
|
|
contract while those on the opposite side of the body relax through |
|||||||||||
Fig. |
15.7. |
Integration |
by |
Interneurons |
|
inhibition. Similarly, when ventral skin is pressed, the ventral longitudinal |
||||||||
muscle contracts, while dorsal muscle on the opposite side of the animal |
||||||||||||||
used in leech bending reflex. When the |
relaxes. (B) Scheme for connections of the four Ρ cells that connect to the |
|||||||||||||
surface of a leech is pressed on one part, |
intemeurons that in turn excite or inhibit motoneurons used for bending. |
|||||||||||||
a reflex bend occurs in the body so as to |
(C) When both Ρ cells are activated, by pressing simultaneously at X and |
|||||||||||||
move that region away from the stimulus. |
at Y, or by pressing at Z, the intemeurons integrate the information and |
|||||||||||||
Touch |
(T) |
and |
pressure |
(P) sensory |
give rise to a vector producing an appropriate bend. The direction of the |
|||||||||
neurons convey |
signals |
to |
an |
array |
of |
bend can be accurately predicted from the firing frequencies of the two Ρ |
||||||||
interneurons. |
|
|
|
|
|
|
|
|
cells- (After Lewis and Kristan, 1998.) |
|
|
|||
Повторная активность и блок проведения сигнала
Угнетение проведения сигнала представляет собой еще один хорошо изученный механизм регулирования работы синапса. В ЦНС пиявок и тараканов, а также в двигательных аксонах ракообразных56) в ответ на серию потенциалов действия физиологической частоты возникает угнетение проводимости сигнала на определенных участках ветвления. В Т, N и Ρ нейронах пиявки механизм подобного ингибирования зависит от гиперполяризации, вызываемой электрогенным натриевым насосом и длительными изменениями проводимости кальций-активируемых калиевых каналов. Так, например, частые повторные прикосновения или давление на кожу пиявки вызывают серию импульсов и длительную гиперполяризацию Ρ клеток. В результате это-
Глава 15. Клеточные механизмы интеграции υ поведения |
325 |
го проведение сигнала блокируется в точках ветвления в области нейропиля, где создаются неблагоприятные условия для проведения импульса34). Именно здесь аксоны малого диаметра переходят в большие по диаметру (глава 7), в то время как в других веточках этого же нейрона сигнал проводится57).
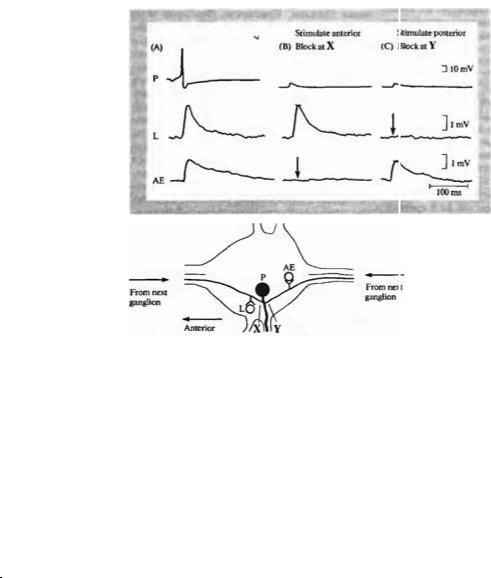
Блок проведения является несинаптическим механизмом, который на время отсоединяет Т или Ρ клетки от некоторых их мишеней. Когда же блокированными оказываются не все, а только часть волокон, освобождение передатчика происходит, но в меньшем количестве. Повреждая при помощи лазера определенные точки ветвления, Мюллер с коллегами сумели оценить вклад отдельных веточек Ρ клеток58). Пример блока проведения в Ρ клетках показан на рис. 15.8. Синаптические потенциалы, записанные на пресинапсах АЕ мотонейронов, полностью исчезли при блоке проведения в ответ на повреждения лазером, нанесенные в точках «X» (рис. 15.8В). Однако одновременно с этим синаптические потенциалы были зарегистрированы в L мотонейроне, иннервируемом той же Ρ клеткой, поскольку этот мотонейрон получал сигналы от веточек, проведение в которых не было нарушено. Повторная активность в физиологическом диапазоне частот привела (в этом случае) к достоверному уменьшению поверхности кожи, с которой вызывались ответы АЕ мотонейрона, т. е. к временному сокращению размеров рецепторного поля57).
Блокирование проведения может привести порой к неожиданному эффекту. Потенциал действия, который не смог перейти с малого аксона на большой, вызывает на последнем локальный потенциал. Этот деполяризационный потенциал, имеющий подпороговую амплитуду, длится гораздо дольше, чем исходный потенциал действия. Таким образом, может возникнуть второй потенциал действия, распространяющийся ретроградно от точки ветвления в сторону терминали чувствительной клетки и там вызывающий повторное выделение медиатора, с небольшой задержкой после первого импульса59).
Блок проведения в дорзальных аксонах спинного мозга млекопитающих впервые был описан Барроном и Метьюсом в 1935 году60), затем прочно забыт и только гораздо позже стал изучаться на беспозвоночных. Сейчас уже очевидно, что блок проведения является ключевым моментом интегративных процессов на дендритах ЦНС млекопитающих61).
Высшие уровни интеграции
Одной из целей изучения таких животных, как пиявки, является исследование того, каким образом простейшие элементарные рефлексы образуют сложные поведенческие реакции. Пиявка оказалась особенно подходящим объектом для изучения путей и отдельных клеток, которые действуют слаженно во время плавания. Этот сложный тип движений изучался Стентом, Кристаном, Фризером и их коллегами. Активность головного и хвостового «мозга» вовсе не являются необходимым условием для осуществления плавательного движения, которое возникает и в нескольких изолированных сегментах и даже всего в одном сегменте. Как и у других беспозвоночных (таких как тараканы, саранча и сверчки), у которых высшие моторные программы с участием небольшого количества нейронов способны контролировать сложные движения, базовый ритм движений пиявки задается рядом возбуждающих и тормозных взаимодействий в ЦНС. Периферические рецепторы служат для регуляции, усиления, ослабления или полной остановки движений пиявки. Сходным же образом препараты изолированного мозга млекопитающих способны генерировать ритм дыхательных движений in vitro (глава 22).
У позвоночных, в свою очередь, интересную роль в регуляции двигательной активности играют биогенные амины62· 63). В крови неподвижных, вялых и не плавающих пиявок уровень серотонина ниже, чем у активных пиявок. Более того, стимуляция секретирующих серотонин клеток (известных как клетки Ретциуса (Retzius cells)), приводит к увеличению его концентрации в крови, и в конечном счете способствует повышению активности всего животного. Можно полностью удалить серотонин из крови эмбриона при помощи специального реактива (5,6-дигидрокситриптамин), который избирательно разрушает серотонин--эргические нейроны в развивающихся ганглиях. Такое взрослое животное само по себе не способно плавать, однако плавательные движения появляются при добавлении в среду серотонина.
Сенситизация и S интернейроны
Механизм того, как отдельный нейрон может играть координирующую роль в поведении животного, тщательным образом изучал-
326 |
|
|
|
|
|
|
|
Раздел III. Интегративные механизмы |
|
Рис. 15.8. Роль блока |
|
|
|
||||||
проведения |
сигнала |
в |
|
|
|
||||
процессах |
синаптической |
|
|
|
|||||
передачи |
и |
интеграции. |
|
|
|
||||
Медиально расположенная |
|
|
|
||||||
Ρ клетка |
(иннервирующая |
|
|
|
|||||
дорзальный |
участок тела, |
|
|
|
|||||
как |
показано |
на |
рис. |
|
|
|
|||
15.4) |
|
|
|
посылает |
|
|
|
||
пресинаптические аксоны к |
|
|
|
||||||
моторным |
|
|
|
клеткам, |
|
|
|
||
вызывающим сокращение |
|
|
|
||||||
продольных |
мышц тела (L |
|
|
|
|||||
клетки) |
и |
образование |
|
|
|
||||
складок (АЕ клетки). (А) |
|
|
|
||||||
Синаптические |
|
|
|
|
|
|
|||
потенциалы в L и АЕ |
|
|
|
||||||
клетках, |
вызванные |
при |
|
|
|
||||
стимуляции |
Ρ |
нейронов |
|
|
|
||||
внутриклеточно |
|
|
|
|
|
|
|||
введенным |
электродом. |
|
|
|
|||||
Импульсы, |
возникшие в |
|
|
|
|||||
соседних сегментах тела в |
|
|
|
||||||
ответ |
|
на |
|
давление, |
|
|
|
||
распространяются |
|
по |
|
|
|
||||
малым аксонам в составе |
|
|
|
||||||
соединительных |
|
веточек |
|
|
|
||||
(коннектив), вовлекают все |
|
|
|
||||||
ветви |
Ρ |
нейрона |
и |
|
|
|
|||
вызывают |
сина |
птические |
|
|
|
||||
потенциалы как на АЕ, так |
|
|
|
||||||
|
|
||||||||
и на L клетках. (В) При |
|
на все веточки Ρ клетки, так же как и в случае А. Однако после многократной |
|
||||||
многократном |
|
|
|
|
стимуляции возникает блок в точке Υ, где аксон малого диаметра переходит в |
||||
раздражении |
|
кожи |
в |
|
|||||
|
|
большой аксон. И вновь это приводит к изоляции клетки (показано стрелкой). |
|||||||
области самого переднего |
|
Однако сейчас изолированной оказывается L клетка, а АЕ клетка остается |
|||||||
сегмента тела, Ρ клетка |
|
активной. |
|||||||
гиперполяри |
зуется, |
и |
|
|
|
||||
проведение |
|
|
сигналов |
|
Fig. 15.8. Effects of Conduction Block on synaptic transmission and integration. The |
||||
блокируется в точке X. При |
|
medially situated Ρ cell (innervating dorsal skin and similar to that shown in Figure |
|||||||
этом L клетка no-прежнему |
|
15.14) sends presynaptic axons to motor cells that cause the longitudinal muscles to |
|||||||
продолжает |
|
|
получать |
|
contract (L cell) and the annul! to erect (AE cell). (A) The synaptic potentials in the L cell |
||||
импульсы, |
|
|
которые |
|
and the AE cell were evoked by stimulating the Ρ cell intracellularly. When the impulses |
||||
вызывают на ее дендритах |
|
originate in the neighboring segment by pressure, they travel along the small axons in |
|||||||
синаптический |
потенциал, |
|
the connective, successfully invade all the branches of the Ρ cell, and produce synaptic |
||||||
однако |
АЕ |
|
клетка |
|
potentials in the AE and the L cell. (B) When the skin in the next most anterior segment |
||||
оказывается |
на |
время |
|
is pressed repeatedly, the Ρ cell becomes hyperpolarized and impulse conduction fails |
|||||
изолированной |
(показано |
|
at the point labeled X. When this occurs, the L cell still receives its input and produces a |
||||||
стрелкой). (С) Наоборот, |
|
synaptic potential but the AE cell has been temporarily disconnected (arrow). (C) |
|||||||
при |
стимуляции |
самого |
|
Conversely, when the posterior field is stimulated, impulses initially invade all the |
|||||
заднего сегмента |
сначала |
|
branches in the Ρ celt as in A. After repetitive stimulation, however, they become |
||||||
импульсы |
|
|
|
|
|
|
blocked at Y, where the small axon encounters the larger axon. Once again cells |
||
распространяются |
|
|
|
become disconnected (arrow). This time, however, it is the L cell that fails to show a |
|||||
|
|
|
|
|
|
|
|
synaptic potential, while the AE cell continues to do so. (After Gu, 1991.) |
|
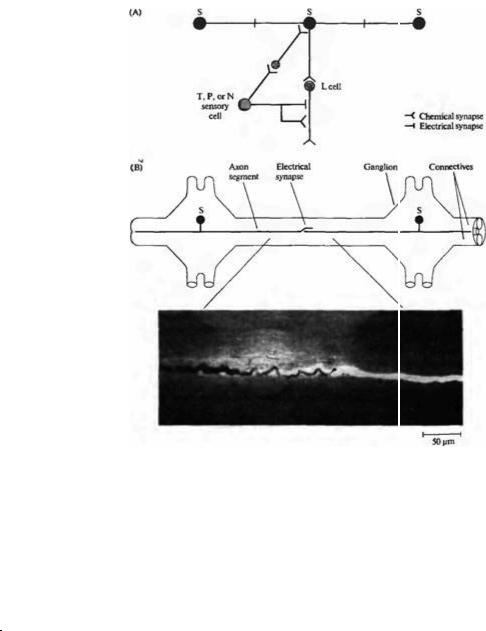
ся на насекомых и ракообразных, у которых были описаны отдельные «командные» нейроны, способные дирижировать оркестром поведенческих реакций всего организма. У пиявки эту роль играет непарный интернейрон, присутствующий в каждом ганглии и называемый S клеткой (рис. 15.2 и 15.9)64). Этот нейрон получает возбуждающие импульсы от чувствительных клеток при касании и давлении и в свою очередь возбуждает L мотонейрон, который способствует сокращению тела (как ранее было указано). Каждая S клетка связана с S клеткой соседних сегментов посредством толстого аксона с большой скоростью проведения через электрический синапс. То, что синапс находится посередине между сегментами (см. рис. 15.9), можно определить путем введения в S клетку пероксидазы хрена, которая не может распространяться через щелевые контакты.
Глава 15. Клеточные механизмы интеграции и поведения |
327 |
||||||||
Рис. |
15.9. |
Электрические |
|
|
|||||
синапсы |
|
с |
|
высокой |
|
|
|||
скоростью |
|
проведения |
|
|
|||||
образуются |
между |
S |
|
|
|||||
клетками |
|
посередине |
|
|
|||||
соединительных |
|
ветвей |
|
|
|||||
(коннектив) |
|
|
|
между |
|
|
|||
сегментами. |
|
|
(А) |
|
|
||||
Электрические |
|
|
|
и |
|
|
|||
химические |
соединения |
|
|
||||||
между |
сенсорными N, |
Т |
|
|
|||||
или Ρ клетками с S клеткой |
|
|
|||||||
и L мотонейроном. В |
|
|
|||||||
каждом |
ганглии |
находится |
|
|
|||||
только одна S клетка, |
|
|
|||||||
аксон |
которой |
образует |
|
|
|||||
электрические |
синапсы |
с |
|
|
|||||
S |
клетками |
|
соседних |
|
|
||||
сегментов. |
|
|
|
|
|
|
|
||
Распространяясь |
|
вдоль |
|
|
|||||
цепочки |
S |
клеток |
по |
|
|
||||
всему |
телу |
животного, |
|
|
|||||
сигналы |
|
чувствительных |
|
|
|||||
клеток способны |
привести |
|
|
||||||
к быстрому |
сокращению |
|
|
||||||
всего тела. |
(В) |
S |
клетки |
|
|
||||
ганглия |
связаны |
с |
S |
|
|
||||
клетками других |
ганглиев |
|
|
||||||
посредством |
|
|
|
|
|
|
|||
электрических |
|
синапсов. |
|
|
|||||
На |
|
микрофотографии |
|
|
|||||
представлена |
S |
клетка, |
|
|
|||||
заполненная смесью люци |
|
|
|||||||
фера |
|
желтого |
|
(Lucifer |
|
|
|||
yetlow) (ЛЖ) и пероксидазы |
|
|
|||||||
хрена |
(horseradish |
peroxi |
|
|
|||||
dase) (ПХ). ПХ неспособна |
|
|
|||||||
проникать |
|
|
|
через |
|
|
|||
электрические |
синапсы, в |
|
|
||||||
отличие от ЛЖ. Поэтому |
|
|
|||||||
соседняя |
|
|
|
клетка |
|
|
|||
окрашивается |
только ЛЖ, |
Fig. 15.9. Connections of S Cells that conduct rapidly and make |
electrical synapses in |
||||||
но не ПХ. Аксон S клетки |
|||||||||
способен |
регенерировать, |
the midregion of the connective. (A) Electrical and chemical connections of sensory T, |
|||||||
восстанавливая |
|
свои |
P, or N cells, and S cells onto the L motoneuron. Each ganglion contains only one S |
||||||
прежние |
|
соединения |
с |
cell, the axons of which make electrical synapses on its homologues. Through the S |
|||||
высокой |
точностью. |
На |
cell chain along the animal sensory stimuli cause rapid shortening. (B) S cells are |
||||||
микрофотографии |
показан |
connected to their homologues in neighboring ganglia by electrical synapses. The |
|||||||
аксон, |
|
|
полностью |
photomicrograph shows an S cell that was filled with a mixture of Lucifer yellow and |
|||||
идентичный |
аксону |
S |
horseradish peroxidase (HRP). HRP cannot cross electrical junctions, whereas Lucifer |
||||||
клетки |
|
|
нормального |
yellow can. Accordingly, the neighboring S cell axon became labeled with Lucifer yellow |
|||||
животного, |
|
|
|
однако |
but not with HRP. S cell axons regenerate and re-form their connections with very high |
||||
фотография была сделана |
specificity after injury. The micrograph, which resembles that of a normal ammaL was in |
||||||||
после того, как данная S |
fact taken from a preparation in which the axon of one S cell had been severed and had |
||||||||
клетка была повреждена и |
re-formed its connections after regeneration. (After Mason and Muller, 1996; |
||||||||
регенерировала, |
|
|
|
micrograph kindly provided by K. Muller.) |
|
||||
|
|
|
|
|
|||||
восстановив свои прежние |
|
|
|||||||
соединения. |
|
|
|
|
|
|
|||
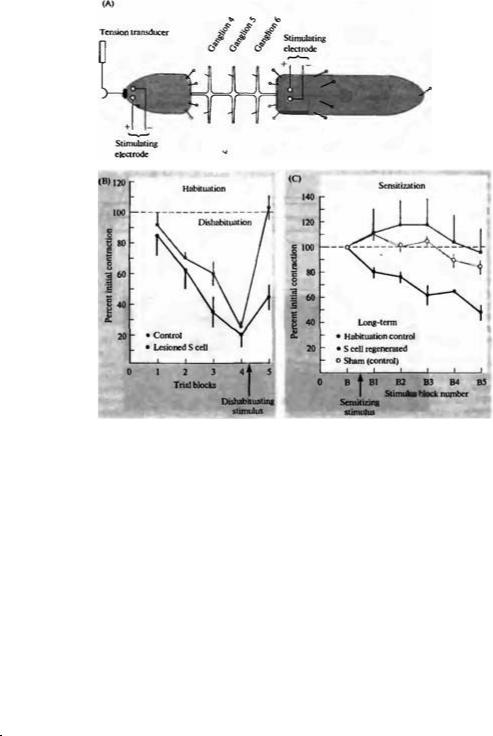
Последовательная цепь S клеток является ключевым механизмом адаптивных реакций пиявки. При повторной тактильной стимуляции прикосновением к определенному сегменту тела сначала возникает рефлекс сокращения. Ответ становится все слабее и слабее при каждом следующем прикосновении — данный эффект известен как «привыкание» (habituation, рис. 15.9)65). Однако после более сильного раздражения, когда активируются Ρ (давление) и N (боль) клетки, ответ на дальнейшую стимуляцию только Т клеток (при касании) вновь становится заметным. Этот процесс известен как «отвыкание» (disha-
328 |
Раздел III. Интегративные механизмы |
Рис. |
|
|
|
|
15.10. |
|
|
|
|
|
|
Процессы |
|
|
|
|
|
|
|
|
|
||
«привыкания» |
|
|
|
|
|
|
|
||||
(habituation), |
|
|
|
|
|
|
|
|
|||
«отвыкания» |
|
|
|
|
|
|
|
|
|||
(dishabituation) |
|
и |
|
|
|
|
|
||||
сенситизации |
|
|
|
|
|
|
|
|
|||
рефлексов у пиявки. |
|
|
|
|
|
||||||
Роль S клеток. (А) |
|
|
|
|
|
||||||
Изолированный |
|
|
|
|
|
|
|||||
ганглий |
соединен |
с |
|
|
|
|
|
||||
передней |
и |
задней |
|
|
|
|
|
||||
частью |
|
|
|
тела. |
|
|
|
|
|
||
Стимулы наносились |
|
|
|
|
|
||||||
на переднюю |
или |
|
|
|
|
|
|||||
заднюю |
часть |
кожи |
|
|
|
|
|
||||
при |
|
|
|
помощи |
|
|
|
|
|
||
микрозлектродов |
|
|
|
|
|
|
|||||
или |
механически, |
|
|
|
|
|
|||||
для активации Т, Ρ |
|
|
|
|
|
||||||
или |
N |
сенсорных |
|
|
|
|
|
||||
нейронов. |
|
|
|
|
|
|
|
|
|
||
Активность S клеток, |
|
|
|
|
|
||||||
сенсорных |
клеток |
и |
|
|
|
|
|
||||
мотонейронов |
|
|
|
|
|
|
|
||||
регистрировалась |
|
|
|
|
|
|
|||||
внутриклеточными |
|
|
|
|
|
|
|||||
электродами, |
|
сила |
|
|
|
|
|
||||
сокращения |
|
мышц |
|
|
|
|
|
||||
определялась |
при |
|
|
|
|
|
|||||
помощи |
|
|
датчика |
|
|
|
|
|
|||
давления. |
|
|
При |
|
|
|
|
|
|||
многократной слабой |
|
|
|
|
|
||||||
стимуляции |
|
|
|
|
|
|
|
|
|||
хвостового |
|
|
конца |
|
|
|
|
|
|||
животного возникало |
|
|
|
|
|
||||||
«привыкание». |
|
|
|
|
|
|
|
||||
Более |
|
|
сильные |
|
|
|
|
|
|||
импульсы |
|
|
|
|
|
|
|
|
|
||
приводили |
|
|
|
к |
линией с |
черными |
кружками). (С) |
После сильного сенситизирующего импульса |
|||
сенситизации |
|
|
и |
||||||||
«отвыканию». |
(В) |
(сенситизация, показано серой линией с серыми кружками) ответы становились выше, |
|||||||||
Ответы |
на |
слабые |
чем обычно при «привыкании». При выключении S клетки либо путем летальной |
||||||||
удары |
током |
кожи |
инъекции проназ, либо при помощи перерезания ее аксона, сенситизация более не |
||||||||
задней |
области. |
В |
выявлялась (черная линия) и многократные импульсы вызывали только «привыкание». |
||||||||
сериях |
стимуляций |
После того, как поврежденная S клетка регенерировала (подобно тому, как показано на |
|||||||||
ответы |
становились |
рис. 15.9), стимул высокой амплитуды вновь вызывал хорошо заметную сенситизацию |
|||||||||
все |
меньше |
|
и |
(серая линия с полыми кружками) и «отвыкание» (не показано на графике). Эти |
|||||||
меньше |
|
|
|
|
|
эксперименты показали, что отдельная S клетка является ключевым звеном в |
|||||
(«привыкание», |
|
|
формировании сложных реакций нервного ганглия. |
||||||||
показано |
|
|
|
серой |
Fig. 15.10. Habituation, Dishabituation, and Sensitization of leech reflexes, and the role of the S |
||||||
линией |
с серыми |
||||||||||
кружками). После |
cell. (A) Isolated ganglia connected to the anterior and posterior parts of the body. Stimuli are |
||||||||||
четырех |
серий |
с |
applied to anterior or posterior skin by electrodes or by mechanical stimuli that activate T, P, or |
||||||||
целью |
|
|
вызвать |
N cells. Intracellular recordings are made from S cells, sensory cells, and motoneurons, and |
|||||||
«отвыкание» |
|
|
|
muscle contractions are measured with a tension transducer. Weak repetitive stimuli applied to |
|||||||
|
|
|
the tail end of the animal give rise to habituation. A stronger stimulus gives rise to sensitization |
||||||||
подавался |
|
|
|
|
|||||||
|
|
|
|
and to dishabituation. (B) Responses to weak electrical shocks to posterior skin. In repeated |
|||||||
высокоамплитудный |
|||||||||||
импульс, |
|
ответ |
на |
trials, |
the |
responses |
became smaller |
(habituation, grey circles). After the fourth triaL a |
|||
который |
|
|
|
был |
dishabituating strong stimulus was given that produced a bigger response. After elimination of |
||||||
значительно |
больше |
the S |
cell |
by killing it in one segment only habituation occurred (black circles). (C) When a |
|||||||
по амплитуде. Когда |
strong |
sensitizing stimulus was given |
(sensitization, grey circles), the responses became |
||||||||
S |
|
|
|
клетка |
stronger than normal instead of habituating. After elimination of the S cell by killing it in one |
||||||
выключалась |
|
|
|
segment or by cutting its axon acutely, sensitization could no longer be elicited (black circles). |
|||||||
|
|
|
Repeated stimuli gave rise only to habituation. After an S cell axon had been severed and then |
||||||||
(инъекцией |
проназ), |
||||||||||
наблюдалось |
только |
regenerated in the manner shown in Figure 15.9, a strong stimulus once again gave rise to |
|||||||||
привыкание |
|
|
|
clear sensitization (grey line with white circles) and dishabituation (not shown). This experiment |
|||||||
|
|
|
demonstrates that a single cell is essential for these complex responses. (After Sahley et al., |
||||||||
(показано черной |
|
||||||||||
|
1994, and Modney, Sahley, and Muller, 1997.) |
||||||||||
|
|
|
|
|
|
|
|||||

Глава 15. Клеточные механизмы интеграции υ поведения |
329 |
Рис. 15.11. Пустынный муравей (Cataglyphis bicolor),
способный ориентироваться по ультрафиолетовому излучению и возвращаться к гнезду кратчайшим путем во время поиска пищи.
Fig. 15.11. The Desert Ant Cataglyphis bicolor. which navigates successfully by means of detection of ultraviolet light. It is able to return home in a direct path after having searched for food. (After Wehner, 1994b.)
bituation) Сходным же образом, если более сильный раздражитель наносится до процедуры «обучения», это вызывает сенситизацию, т. е. ответ на стимуляцию при касании будет больше обычного. Активность S клеток во время «привыкания» не меняется, однако возрастает как при сенситизации, так и при «отвыкании» (см. рис. 15.10). В технически сложно выполнимых экспериментах, когда избирательно перерезается аксон S клетки, либо клетки убиваются инъекцией проназ, процессы «привыкания» и сокращения не изменяются, однако полностью исчезает «отвыкание» и сенситизация.
Во второй серии экспериментов аксоны S клеток были повреждены, чтобы исследовать их регенерацию. Удивительным свойством S клеток является то, что после повреждения их аксоны следуют по старому пути и заново образуют электрические соединения с соседним S нейроном с поразительной точностью66). Как и ожидалось, перерыв передачи импульсов по S клеткам между отдельными сегментами приводил к исчезновению сенситизации, однако через несколько недель после полной регенерации аксонов и восстановления прежних связей данное свойство также восстанавливалось67).
Эти примеры наглядно демонстрируют, каким образом отдельная нервная клетка способна принимать участие в таком сложном поведенческом процессе, как сенситизация. Было установлено, что на самом деле процессы сенситизации во многом обусловлены серотонином68). (Необычная серия совпадений способствовала тому, что название «S клетка» стало очень подходящим для данных клеток. Изначально она была так названа, потому что ее импульс образует долгий импульс (spike) в месте контакта. Затем, обнаружив, что эти клетки очень тесно связаны друг с другом, полагали, что они могут быть синцитием. Сейчас же они называются S клетками от слов «сенситизация» и «серотонин»!) У Aplysia процесс сенситизации (в котором серотонин тоже играет важную роль) был очень тщательно изучен как на клеточном, так и на молекулярном уровне69).
§ 3. Навигация у пчел и муравьев
Одним из основных методов исследования функционирования НС является количественный анализ поведения. При анализе поведения можно выявить фундаментальные принципы. Например, ключевые механизмы работы зрительной системы, которые смогли объяснить цветовое зрение и темновую адаптацию у человека, были впервые установлены при помощи психофизических экспериментов. При помощи изучения реакции разных людей на световые сигналы различной интенсивности и длины волны стало возможным сделать вывод о механизмах светового восприятия. Только значительно позже клеточные механизмы были показаны на фоторецепторах и зрительных путях (главы 19 и 20). Аналогичным образом, наблюдение за беспозвоночными в естественных условиях привело к ценным открытиям роли рецепторов и описанию интегративных функций ЦНС.
Удивительные характеристики НС беспозвоночных вызывают восхищение, например, при изучении процессов навигации у пчел и муравьев70· 71). Пустынный муравей может мигрировать, а пчела улетать на далекие расстояния от гнезда в поисках пищи, и затем чудесным образом они безошибочно находят дорогу обратно. Необходим целый набор различных сенсорных «подсказок» для того, чтобы подобная навигация стала возможной. Изза того, что нервные клетки насекомых имеют маленькие размеры, зачастую довольно сложно непосредственно анализировать свойства их мембран и синаптическую передачу. Тем не менее, как будет показано в следующих разделах, можно проанализировать и сделать определенные выводы о сенсорных механизмах насекомых даже на клеточном уровне при

330 |
|
|
|
|
Раздел III. Интегративные механизмы |
||||
Рис. 15.12. Для изучения перемещения муравья |
|
|
|||||||
на поверхности пустыни была нанесена сетка. (А) |
|
|
|||||||
Полевые эксперименты по прослеживанию пути |
|
|
|||||||
пустынного муравья, выполняемые при помощи |
|
|
|||||||
специальной оптической системы на роликах, |
|
|
|||||||
способной следовать эа муравьем во время его |
|
|
|||||||
движения и регистрировать траекторию его |
|
|
|||||||
перемещений, а также контролировать и |
|
|
|||||||
манипулировать углом обзора и оптическими |
|
|
|||||||
свойствами |
воспринимаемых |
|
муравьем |
|
|
||||
солнечных лучей. (В) Камера перемещалась |
|
|
|||||||
экспериментатором таким образом, чтобы |
|
|
|||||||
муравей всегда находился в оптическом центре |
|
|
|||||||
системы. В горизонтальную апертуру (1) |
|
|
|||||||
устанавливались |
|
различные |
|
фильтры, |
|
|
|||
отсекающие |
излучение |
за |
|
пределами |
|
|
|||
ультрафиолетового спектра |
или |
пропускающие |
|
|
|||||
лучи только определенной поляризации. Малая |
|
|
|||||||
апертура (2), расположенная на цилиндре, а |
|
|
|||||||
также |
экран |
(3) |
использовались |
для |
|
|
|||
предупреждения |
падения |
прямого |
солнечного |
|
|
||||
света на фоторецепторы муравья. Из-за высокой |
|
|
|||||||
боковой рамы камеры муравей не мог видеть |
|
|
|||||||
линию горизонта или какие--либо метки на |
|
|
|||||||
поверхности пустыни. Это также ограждало его от |
|
|
|||||||
ветра. Линии белого цвета, нанесенные на |
|
|
|||||||
расстоянии 1 м друг от друга на поверхности |
|
|
|||||||
пустыни, позволяли экспериментатору тщательно |
|
|
|||||||
следить за перемещениями муравья. |
|
|
|
|
|||||
Fig. 15.12. То Measure the Movements of an Ant it is |
|
|
|||||||
placed on a grid marked out on the desert floor (A). A |
|
|
|||||||
desert ant is tracked in the field by manipulating its |
|
|
|||||||
skylight vision experimentally and simultaneously |
|
|
|||||||
recording its walking trajectory using a rolling optical |
|
|
|||||||
laboratory for following ants as they migrate along the |
|
|
|||||||
desert floor and for controlling the portion of sky that |
|
|
|||||||
they can observe (B). The trolley is moved by the |
|
|
|||||||
experimenter in such a way as to keep the ant always |
|
|
|||||||
centered within the optical setup. The horizontal |
|
|
|||||||
aperture (1) can be fitted with filters that cut out all |
|
|
|||||||
light apart from ultraviolet or that allow polarization in |
|
|
|||||||
one direction. A small aperture (2) sits atop a circular |
It is also shielded from the wind. The white lines are 1 m |
||||||||
tube. and the screen (3) can be used to prevent the |
apart and |
are painted on the desert floor to enable the |
|||||||
ant from seeing sunlight directly. Since the trolley has |
observers |
to track the anf s progress accurately. (After |
|||||||
a frame, the little ant cannot see |
the |
skyline or |
Wehner, 1994b, photograph kindly provided by R. Wehner.) |
||||||
markers on the ground. |
|
|
|
|
|
|
|
||
помощи особым образом спланированных поведенческих экспериментов.
Как пустынный муравей находит дорогу домой
Венер с коллегами экспериментально изучал, каким образом пустынный муравей Cataglyphis bicolor (рис. 15.11) способен мигрировать на далекие дистанции в поисках пищи и затем возвращаться к гнезду по прямому кратчайшему пути. Принцип их экспериментов показан на рис. 15.12 и 15.1372· 73). Область вокруг гнезда (в пустыне Туниса), а также места расположения пищи отмечены квадратиками. За отдельным муравьем наблюдали во время того, как он искал источник пищи и возвращался с ней в гнездо. На рис. 15.13 длинный и извилистый путь от гнезда (от пункта А до пункта В) занимает около 19 минут (точка на маршруте равна 1 минуте). Дорога домой, наоборот, идет строго по прямой, не петляет и занимает около 6 минут. Каким-то образом муравей учел свои движения на протяжении 250 метров от гнезда. Каким--то образом он также отследил углы поворотов и пройденные дистанции. Муравей способен отойти от гнезда на расстояние более 100 метров и вернуться к нему обратно с точность до 1 метра, т. е. с ошибкой менее 1 % (в этом отношении муравей способен сделать это гораздо лучше, чем по крайней мере
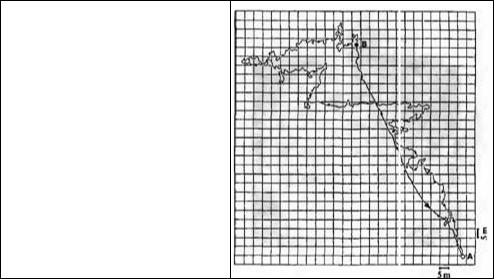
Глава 15. Клеточные механизмы интеграции υ поведения |
331 |
Рис. 15.13. Путь, пройденный муравьем от гнезда (А) до источника пищи (В). Каждая точка на пути туда и обратно отмечает промежуток времени в 1 минуту. В поисках пищи муравей проходит извилистый путь длиной 592 метра, однако обратно он следует по кратчайшему пути (140 м), без отклонений и удивительно точно.
Fig. 15.13. Pathway Taken by an Ant from its nest (A) to the source of food (B). Each dot on the trace toward food and back represents 1 minute of time. The ant progresses to the food in a tortuous path over
592 m. But it heads straight for home (140 m) with no deviation and amazing accuracy. (After Wehner,
1994a.)
один из авторов этой книги, который не имеет математических навыков в подобного рода расчетах).
Какие же ориентиры окружающей среды муравей использует для навигации? Первая мысль — может быть, это феромоны или химические сигналы. Однако они не используются и в любом случае были бы бесполезны при очень высокой (до 45° С) температуре в пустыне74). Пространственные ориентиры и характерные элементы ландшафта в пустыне немногочисленны. Хотя предметы по дороге и около гнезда и в самом деле могут давать определенную информацию (как мы очень скоро увидим), они вовсе не являются теми удивительными «компасами», которые используют муравьи. Это было установлено при помощи экспериментов, когда муравьи были способны найти обратный путь, видя всего лишь небольшую часть неба71). Чтобы исключить солнце и все другие ориентиры, экспериментатор следовал за муравьем с небольшой камерой (см. рис. 15.12), держа ее отверстие (через которое было видно небо) точно над муравьем. В отверстие помешались специальные фильтры для выбора направления, длины волны и угла поляризации солнечного света, воспринимаемого муравьем. Даже без солнца, отличительных деталей ландшафта и запахов, муравей, видя только небо, направлялся прямо к гнезду, в то время как экспериментатор катил камеру за ним, скрытый в маленьком вагончике.
Использование поляризованного света как компаса
О поляризации электромагнитного излучения говорят, когда вектор электрической напряженности электромагнитной волны колеблется в одной плоскости. Известно, что солнечный свет становится поляризованным при прохождении атмосферы: через ориентированный определенным образом поляризующий фильтр небо выглядит темным (так как отсекается поляризованный свет). Свет, отраженный от неровной поверхности облаков, однако, уже не является поляризованным. Следовательно, через подобный фильтр облака будут выглядеть более яркими. Когда солнце находится точно в зените, картина поляризации довольно проста. Все направления поляризации в горизонтальной плоскости будут представлены пропорционально, следовательно, это не может служить компасом. Однако, если лучи солнца падают не под прямым углом, возникает асимметричное множество направлений поляризации, как показано на рис. 15.14. Эта картина поляризации может, в принципе, служить картой для навигации. И хотя глаз человека не способен воспринимать поляризованный свет, глаз пустынного муравья на это способен, как, впрочем, и глаза пчел, ос и ракообразных75). Картина поляризации света показывает (определяет) положение солнца вне зависимости от того, можно ли его увидеть напрямую или нет.

332 |
Раздел III. Интегративные механизмы |
Рис. |
15.14. |
Структура |
|
|
|
||||
поляризованного |
|
|
|
|
|
||||
солнечного |
|
света |
и |
|
|
|
|||
регистрация |
его |
при |
|
|
|
||||
помощи |
набора |
разных |
|
|
|
||||
поляризующих |
фильтров. |
|
|
|
|||||
(А, |
|
В) |
|
Набор |
|
|
|
||
поляризующих |
фильтров, |
|
|
|
|||||
установленных |
|
в |
виде |
|
|
|
|||
купола |
|
с |
|
целью |
|
|
|
||
регистрации |
|
картины |
|
|
|
||||
поляризованного |
|
|
|
|
|
||||
солнечного |
света. |
Для |
|
|
|
||||
человека, |
стоящего |
на |
|
|
|
||||
широте |
|
экватора |
в |
|
|
|
|||
полдень, |
когда |
солнце |
|
|
|
||||
|
|
||||||||
находится строго в зените, |
|
Муравью достаточно видеть небольшой кусочек неба, чтобы рассчитать |
|
||||||
картина |
|
|
|
выглядит |
|
|
|
||
|
|
|
положение солнца и правильно сориентироваться. |
||||||
полностью |
симметричной. |
|
|
|
|||||
(С |
D) |
При |
перемещении |
Fig. 15.14. Patterns of Polarized Light and Their Detection by an array of polarizing |
|||||
солнца |
|
из |
|
одного |
|
filters. (A, B) The arrangement of polarizing filters mounted in a dome so as to detect |
|||
положения |
|
в |
другое |
|
patterns of polarized light in the sky. For a person standing at the equator at noon with |
||||
меняется |
|
|
|
и |
|
the sun shining directly overhead, the patterns would be completely symmetrical. (C, D) |
|||
поляризационная |
картина, |
|
The patterns produced by light from the sun at two different positions, indicated by the |
||||||
что |
указано |
|
черными |
|
black dots. The ant need only observe a small part of the heavens in order to compute |
||||
точками. |
|
|
|
|
|
the position of the sun and thereby navigate successfully. (A and В after Wehner, |
|||
|
|
|
|
|
|
|
|
1994a; С and D after Wehner, 1997.) |
|
Восприятие поляризованного света глазом муравья
Мультифасеточный сложный глаз муравья состоит из набора отдельных единиц, омматидий (ommatidia), от каждой из которых идет свое нервное волокно. В каждой омматидии находится 8 фоторецепторов. Каждая омматидия видит мир в собственной перспективе76). У пустынного муравья определенные группы омматидии чувствительны к поляризованному свету строго определенной ориентации. Они расположены в области дорзального ободка (dorsal rim) сложного глаза и воспринимают свет преимущественно в УФ диапазоне. На рис. 15.15 и 15.16 показано расположение этих рецепторов. Их чувствительность к поляризации возникает из-за точной ориентации мембран, содержащих фотопигмент родопсин. Молекулы родопсина расположены параллельно, в однотипно расположенных микроворсинках фоторецепторных клеток, как показано на рис. 15.1677). Так как родопсин оптимально поглощает свет, направленный вдоль длинной оси молекулы, только одна определенная плоскость поляризованно-