

Nikolls_-_Ot_neyrona_k_mozgu
.pdfГлава 12. Синоптическая пластичность
Эффективность синаптической передачи не фиксирована и может изменяться как следствие изменения паттерна текущей активности. Короткие залпы потенциалов действия в пресинаптическом нейроне могут вызвать либо облегчение (фасилитацию) выделения медиатора из пресинаптической терминали на несколько сотен миллисекунд, либо подавление выделения медиатора на секунды, либо комбинацию этих процессов. Вторая фаза фасилитации, называемая усиление (augmentation), также может длиться секунды. Длинные залпы потенциалов действия в пресинаптическом нейроне могут вызвать посттетаническую потенциацию (ПТП), которая характеризуется увеличением выделения медиатора на десятки минут. Увеличение концентрации кальция в пресинаптической терминали лежит в основе этих изменений выделения медиатора.
Во многих синапсах повторяющаяся активность может вызвать не только кратковременные, но и долговременные изменения синаптической эффективности, которые длятся часы и даже дни. Два явления этого рода известны как долговременная потенциация (ДВП) и долговременная депрессия (ДВД). ДВП опосредована увеличением концентрации кальция в постсинаптической клетке, которое запускает каскады систем вторичных посредников (мессенджеров), активность которых приводит к появлению дополнительных рецепторов в постсинаптической мембране и увеличивает чувствительность рецепторов. ДВД появляется в ответ на сравнительно меньшее увеличение концентрации кальция в постсинаптической клетке и сопровождается уменьшением количества и чувствительности постсинаптических рецепторов.
Некоторые формы ДВП и ДВД вовлекают пресинаптические механизмы. Обе эти формы пластичности считаются основой различных форм обучения и формирования памяти, но имеющиеся в настоящее время данные пока не предоставили безоговорочных доказательств этой гипотезы.
До настоящего времени мы обсуждали передачу возбудительных и тормозных сигналов в активированных синапсах в терминах одиночного потенциала действия, приходящего по пресинаптической терминали, деполяризации терминали, входа кальция и высвобождения медиатора, приводящего к деили гиперполяризации постсинаптической мембраны в зависимости от свойств постсинаптических рецепторов. В этих условиях ответы каждого данного синапса относительно стереотипны и постоянны, если не считать естественную вариабельность. Однако при нормальной работе мозга синапсы активируются не отдельными потенциалами действия (ПД), а непрерывными потоками ПД разной частоты и регулярности.
Такая текущая активность может оказывать эффективное воздействие на синаптическую передачу. Например, при стимуляции пресинаптического волокна коротким залпом стимулов результирующие постсинаптические потенциалы могут либо увеличиваться (синаптическая фасилитация), либо уменьшаться (синаптическая депрессия). Эти изменения сохраняются и после окончания вызвавшей их активности, и их классифицируют в соответствии с длительностью сохранения (рис. 12.1). Фасилитация появляется мгновенно и длится все время стимуляции. После того, как залповая стимуляция заканчивается, ответы на одиночные тестовые стимулы показывают, что увеличение синаптической эффективности длится несколько сотен миллисекунд. Более длительная фаза фасилитации, длящаяся несколько секунд, называется усилением ответа (augmentation).
В тех случаях, когда активность синапсов приводит к депрессии, восстановление происходит также за несколько секунд. Относительно длительный, высокочастотный залп стимулов (обычно называемый тетанусом, потому что при приложении такого залпа стимулов к мышце или моторному нерву происходит

244 |
Раздел II. Передача информации в нервной системе |
Рис. 12.1. Динамика изменений эффективности синаптической передачи, вызванная ритмической активацией. (А) фасилитация; (В) усиление и депрессия; (С) ПТП; (D) ДВП.
Fig. 12.1. Time Courses of Activity-Induced Changes in synaptic transmission. (A) The main component of facilitation decays over a period of about 100 ms, with a smaller phase persisting for several hundred ms. (B)
Recovery from depression is complete, and augmentation is largely dissipated, by 10 s. (C) FTP lasts for more than 10 min, and (D) LTP and LTD last well beyond 10 h.
тетаническое мышечное сокращение) обычно приводит к синаптической депрессии, вслед за которой через несколько секунд наблюдается увеличение амплитуды синаптического потенциала, которое может длиться десятки минут. Это называется посттетанической потенциацией (ПТП). Ритмичная активация синапсов в центральной нервной системе может вызывать еще более длительные изменения в синаптической передаче, называемые
долговременной потенциацией (ДВП) и долговременной депрессией (ДВД). Эти явления могут длиться многие часы и даже дни.
При исследовании механизмов, лежащих в основе долговременных изменений эффективности синапсов, прослеживается общая нить. Существенная часть регуляции синаптической передачи опосредована изменениями концентрации внутриклеточного кальция в пресинаптической терминали и/или постсинаптической клетке. Таким образом, синаптическая регуляция существенно зависит от механизмов прохождения ионов кальция через плазматическую мембрану и их мобилизации из внутриклеточных депо. Эти механизмы, которые включают как прямую активацию кальциевых каналов (глава 9), так и сложные системы вторичных посредников (глава 10), являются средством регуляции синаптической эффективности во временных рамках от миллисекунд до часов.
§ 1. Кратковременные изменения
Кратковременные изменения эффективности синаптической передачи наиболее интенсив-
Глава 12. Синаптическая пластичность |
245 |
но были изучены в синапсах периферической нервной системы, таких как синапсы скелетных мышц или автономных ганглиев. Тем не менее, подобные изменения были показаны и практически во всех отделах центральной нервной системы. В настоящем разделе мы обсудим фасилитацию, усиление, депрессию и ДВП нервно-мышечного синапса лягушки и цилиарного ганглия цыпленка. Эти изменения «кратковременные», так как их длительность варьирует от десятков миллисекунд (фасилитация) до десятков минут (ДВП). В особых условиях на этих экспериментальных моделях ДВП может длиться часами.
Фасилитация и депрессия выброса медиатора
Во многих синапсах наиболее быстрый эффект ритмической стимуляции заключается в фасилитации ответа. Это проиллюстрировано на рис. 12.2, на котором показаны потенциалы концевой пластинки нервно-мышечного соединения лягушки, вызванные коротким залпом импульсов, приходящим по моторному нерву. Амплитуда потенциалов (измеренная от начальной точки переднего фронта) увеличивается во время залпа. Более того, эффект наблюдается существенно дольше, чем длится залп стимулов, так что ответ на тестовый стимул через 230 мс все еще больше, чем первый ответ в последовательности. Фасилитация может быть разделена на два неравных компонента: больший компонент уменьшается с постоянной времени около 50 мс, тогда как второй компонент уменьшается с постоянной времени около 250 мс1). Показано, что фасилитация происходит из-за увеличения среднего количества квантов медиатора (m; см. гл. 11), высвобождаемого в пресинаптическом окончании2)--4). Дальнейший статистический анализ выброса медиатора в нервно-мышечном соединении рака привел к предположению, что увеличение m происходит скорее благодаря увеличению р, которое отражает вероятность выброса медиатора, чем из-за увеличения n — количества готовых к выбросу квантов медиатора5). Количество медиатора, высвобождаемое из терминали, может удваиваться при фасилитации. Синаптическая депрессия может наблюдаться в тех случаях, когда количество квантов медиатора, высвобождаемых залпом стимулов, велико. В эксперименте, показанном на рис. Ι2.2Α, амплитуда потенциалов концевой пластинки была уменьшена понижением концентрации кальция в омывающем растворе; в этом случае начальный квантовый состав потенциалов был небольшим (10 или менее). Похожий эксперимент при большой концентрации кальция показан на рис. 12.2Β. В этом эксперименте квантовый выброс значительно больше, но ответы были уменьшены по амплитуде блокадой постсинаптических АХ рецепторов при помощи кураре. При ритмической стимуляции ответы становились все меньше по амплитуде. Как и в случае с фасилитацией, депрессия длится дольше залпа стимулов.
Депрессия потенциала концевой пластинки, как и фасилитация, имеет пресинаптическую природу. Ее механизм не полностью ясен, но тот факт, что для депрессии необходима исходно большая величина квантового выброса медиатора, предполагает, что истощение количества пузырьков в пресинаптической терминали во время залпов стимулов может быть одним из определяющих факторов депрессии6). Вторым фактором может являться модуляция выброса АХ из пресинаптической терминали совместным выбросом АТФ (глава 13). АТФ гидролизуется до аденозина, который затем влияет на пресинаптическую терминаль и уменьшает квантовый состав ответа7). АТФ и сам может угнетать секрецию медиатора через собственные пресинаптические Р2 рецепторы (Giniatullin and Sokolova, 1998). После длительного периода ритмической стимуляции депрессия может быть очень сильной, уменьшая амплитуду синаптического потенциала более, чем на 80 %.
В заключение можно сказать, что выброс медиатора из пресинаптических терминален может претерпевать два типа относительно кратковременных модификаций. Первая, фасилитация, происходит благодаря увеличению эффективности выброса квантов из пресинаптической терминали. Вторая, депрессия, может быть связана как с истощением количества квантов, так и с уменьшением эффективности выброса. На рис. 12.2С проиллюстрировано взаимодействие этих двух эффектов. Во время залпов импульсов начальный эффект фасилитации преобладает над депрессией, и синаптический ответ растет по амплитуде. Позднее, когда дается тестовый стимул, фасилитация уже ослабла, и начинает преобладать депрессия, которая длится дольше.

246 |
Раздел II. Передача информации в нервной системе |
Рис. 12.2. Фасилитация и депрессия в нервно-мышечном синапсе позвоночных.
Fig. 12.2. Facilitation and Depression at the vertebrate neuromuscular junction. (A) Muscle bathed in low calcium solution to reduce the quantum content of the response.
Amplitudes of the end-plate potentials increase progressively during a train of four impulses. The response to a test pulse 230 ms later is still facilitated (arrows indicate initial amplitude). (B) Similar experiment with a curarized preparation in high calcium solution. The response amplitudes decrease progressively during the train. (C) Interaction between facilitation and depression in normal calcium concentration. There is no further facilitation after the second response because of the onset of depression, and the test response at 230 ms is depressed. (A and С after Mallart and Martin, 1968; В after Lundberg and Quilisch, 1953.)
Роль кальция в фасилитации
Данные, полученные Катцем и Миледи8), позволяют предположить, что фасилитация выброса медиатора на второй из парных потенциалов действия (ПД) связана с остаточным внутриклеточным кальцием, вызванным предшествующим ПД. В ряде работ были исключены некоторые другие возможные механизмы этого феномена. Например, в синапсах цилиарного ганглия цыпленка было показано, что фасилитация не является следствием увеличения амплитуды или длительности второго ПД в пресинаптической терминали9). Показано также, что фасилитация второго ответа в синапсах между нейронами пиявки происходит без увеличения входа кальция в пресинаптическую терминаль10). Таким образом, наиболее вероятной является гипотеза о том, что фасилитация связана с остаточным кальцием в терминали. При ритмической стимуляции накопление внутриклеточного кальция приводит к прогрессивному увеличению выброса медиатора, как показано на рис. 12.2А. Некоторые теоретические подходы, используемые при исследовании связи между кинетикой концентрации внутриклеточных ионов кальция и динамикой фасилитации, можно найти в обзоре11).
Усиление синаптической передачи
Охарактеризовав временное течение фасилитации и депрессии, Маглеби и Зенгель обнаружили дополнительный эффект ритмической стимуляции, который они назвали усилением (augmentation)12· 13). Усиление представляет собой вызванное ритмической стимуляцией увеличение амплитуды синаптического потенциала, которое развивается значительно медленнее, чем фасилитация, и спадает за более длительный период с постоянной времени от 5 до 10 с (рис. 12.1В). Как фасилитация, так и усиление происходят из-за увеличения выброса медиатора из пресинаптической терминали. В нервно-мышечном соединении лягушки усиление и фасилитация вместе могут увеличить амплитуду синаптического потенциала более чем в пять раз.
Посттетаническая потенциация
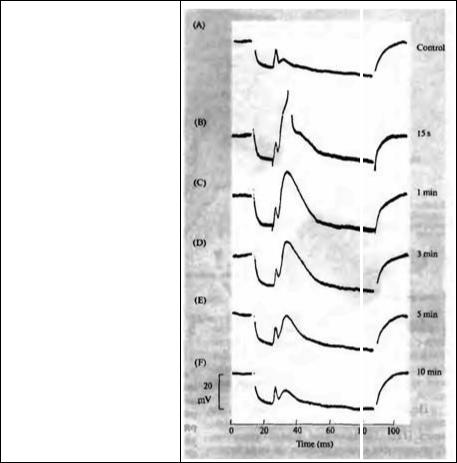
ПТП подобна фасилитации и усилению в том, что она отражает увеличение выброса из пресинаптической терминали, вызванное предшествующей стимуляцией. Различие состоит в том, что ПТП начинается существенно позже, достигает максимума через несколько секунд после конца стимуляции и длится десятки минут. На рис. 12.3 показан пример эксперимента на клетке цилиарного ганглия цыпленка, обработанной кураре для уменьшения амплитуды ВПСП. Кроме того, для предотвращения запуска ПД клетка была гиперполяризована (длинные отклонения вниз на рис. 12.3А) перед стимуляцией пресинапти-
Глава 12. Синоптическая пластичность |
247 |
Рис. 12.3. Посттетаническая потенциация возбудительного постсинаптического потенциала (ВПСП) в клетке цилиарного ганглия цыпленка, вызванная стимуляцией нерва. (А) контрольная запись до потенциации; (В) ответ через 15 с после залпа в 1 500 стимулов; (C-F) ответы на тест--стимулы через 1, 3, 5 и 10 мин после тетанизации.
Fig. 12.3. Posttetanic Potentiation of the excitatory postsynaptic potential (EPSP) in a chick ciliary ganglion cell, produced by preganglionic nerve stimulation. Potentials were recorded with an intracellular microelectrode. To prevent action potential initiation, the EPSP amplitude was reduced with curare, and a hyperpolarizing pulse was applied through the recording electrode before each stimulus. (A) The control record shows electrical coupling potential (brief depolarization) followed by a small EPSP.
(B) The response recorded 15 s after the end of a train of 1 500 stimuli applied to the preganglionic nerve. The EPSP amplitude is more than six times greater than the control, giving rise to an action potential. The amplitude of the coupling potential is unchanged. (C-F) Test stimuli at 1, 3, 5, and 10 rnin after the tetanus show the slow decline of potentiation, with the EPSP in the last record still more than twice the control value. (From Martin and
Pilar, 1964.)
ческого нейрона. Первое отклонение вверх на каждой записи представляет собой потенциал электрической связи между стимулируемой и регистрируемой клеткой; более медленная деполяризация представляет собой ВПСП, вызванный выбросом АХ из пресинаптической терминали (глава 9). В данном случае анализу подвергается ВПСП. Изначально амплитуда ВПСП была только около 4 мВ (из-за присутствия кураре). Затем пресинаптический нерв раздражался с частотой 100 импульсов в секунду в течение 15 с, что вызвало кратковременное уменьшение ВПСП (не показано). Через 15 секунд одиночный стимул вызвал ВПСП амплитудой более 20 мВ, который превысил порог и вызвал ПД (рис. 12.3В). Ответы на последующие стимулы постепенно уменьшались (рис. 12.3C-F), но даже через 10 минут после конца тетанической стимуляции ответ был вдвое больше начального.
Как и фасилитация, ПТП имеет пресинаптическое происхождение. Это означает, что ПТП вызвана увеличенным выбросом медиатора из пресинаптической терминали, скорее всего, из-за увеличения концентрации внутриклеточного кальция. Точный механизм этого явления неясен, но эксперименты на нервно-мышечном соединении лягушки показали, что ПТП зависит от входа кальция в терминаль во время залпа стимулов. Например, если убрать кальций из омывающего раствора во время стимуляции, то потенциация не будет наблюдаться14). В отличие от кальция, вход натрия не является необходимым, так как ПТП может быть вызвана аппликацией деполяризационных импульсов на терминаль в присутствии тетродотоксина15). В этих

248 |
Раздел II. Передача информации в нервной системе |
Рис. 12.4. Гиппокампаль ная формация.
Fig. 12.4. The Hippocampal Formation Lies buried in the temporal Lobe and consists of two interlocking
C-shaped strips of cortex, the dentate gyrus and the hippocampus, together with the neighboring subiculum. Pyramidal cells in area CA1 are innervated by the Schaf-fer collateralcommissural pathway. Granule cells in the dentate gyrus are innervated by the perforant fiber pathway from the subiculum and, in turn, innervate CA3 cells.
условиях величина потенциации увеличивается с увеличением концентрации экстраклеточного кальция. При очень высоких концентрациях кальция (83 ммоль) потенциация после 500 стимулов длится более двух часов.
Несмотря на то, что натрий не нужен для потенциации, его вход влияет на длительность ПТП. Потенциация нервно-мышечного соединения крысы удлиняется воздействиями, которые блокируют выведение натрия Na-K АТФазой, например, добавлением уабаина или удалением калия из омывающего раствора16). Увеличение длительности ПТП может происходить из-за того, что увеличенная концентрация внутриклеточного натрия уменьшает скорость выведения накопленного кальция Na—Са обменом. ПТП в нервно-- мышечном соединении рака также уменьшается по величине и длительности при воздействиях, которые влияют на обмен кальция между цитоплазмой и внутриклеточными хранилищами, например, митохондриями17). Эти эксперименты позволяют предположить, что вход кальция во время тетануса приводит к быстрому наполнению внутриклеточных хранилищ кальция. Накопленный кальций после тетануса медленно высвобождается в цитоплазму, поддерживая повышенную концентрацию свободного кальция в цитоплазме.
§ 2. Долговременные изменения
Ритмическая активность нейронов центральной нервной системы может вызывать изменения синаптической эффективности, которые длятся значительно дольше, чем в периферических синапсах. Длительные изменения могут наблюдаться в разных частях мозга, и это явление особенно интересно тем, что может быть связано с памятью.
Долговременная потенциация
Долговременная потенциация (ДВП) впервые была описана Блисс, Ломо в 1973 году в глутаматергических синапсах гиппокампальной формации18). Эта структура находится в височной области мозга и состоит из двух областей, известных как гиппокамп и зубчатая фасция, которые на срезе видны как две полоски коры С-образной формы, взаимно охватывающие друг друга и субикулума (рис. 12.4). Упорядоченное расположение клеток и входных путей позволяет стереотаксически ввести в мозг животного электроды точно к определенным типам клеток и внутриклеточно регистрировать синаптические потенциалы. Соответственно, легко расположить и стимулирующие электроды на специфических входных путях.

Глава 12. Синоптическая пластичность |
249 |
Рис. 12.5. Долговременная потенциа ция (ДВП) в гиппокампе анестезированного кролика. (А) Синаптические ответы на стимуляцию перфорантного пути регистрировались
вгранулярных клетках зубчатой фасции. (В) Короткие тетанусы (15 ст/с
втеч. 10 с) наносятся в отмеченные стрелками моменты.
Fig. 12.5. Long-Term Potentiation (LTP) in the hippocampus of an anesthetized rabbit. (A) Synaptic responses to perforant fiber pathway stimulation were recorded from granule cells in the dentate gyrus. (B) Brief tetanic stimuli (15/s for 10 s) were given at times marked by the arrows. Each tetanus caused an increase in the amplitude of the synaptic response (grey circles), eventually lasting for hours. In the same cell, responses to stimulation of a control pathway not receiving tetanic stimulation (black circles) were unchanged. (After Bliss and L0mo, 1973.)
Блисс и Ломо показали значительное увеличение синаптических входов клеток зубчатой фасции в результате высокочастотной стимуляции входов к этим клеткам, которое длится часы
идаже дни (рис. 12.5). Сейчас это явление известно как гомосинаптическая ДВП. Несмотря на то, что ДВП была показана и в других областях мозга, включая несколько отделов новой коры,
идаже в нейромышечном соединении ракообразных19), наиболее интенсивно это явление было изучено на срезах гиппокампа 20· 21).
Ассоциативная ДВП в пирамидных клетках гиппокампа
Браун с соавторами показали, что для возникновения ДВП ассоциированная во времени слабая стимуляция двух входов эффективнее раздельной стимуляции22). Это явление названо ассоциативной ДВП. Пример этого явления показан на рис. 12.6. Внутриклеточно отводилась активность пирамидных клеток области СА1 гиппокампа. Два экстраклеточных стимулирующих электрода были расположены на входных путях (коллатерали Шаффера) таким образом, чтобы активировать разные субпопуляции аксонов, активирующих дендритное дерево пирамидной клетки (рис. 12.6А). Интенсивность стимуляции была подобрана таким образом, что электрод I вызывал большой синаптический потенциал в пирамидной клетке, тогда как электрод II существенно меньший.
Синаптические ответы на стимуляцию через электрод II показаны на рис. 12.6В. Короткий залп импульсов (100 Гц в течение 1 секунды, повторенный через 5 с) через электрод I привел к усилению ответов, активируемых через этот вход (не показано). Тетанизация, примененная через электрод I, не оказала влияния на величину ответа, вызываемого входом II (рис. 12.6В, второй ответ). Кроме того, высокочастотная тетанизация через вход II также не вызвала изменений тестового ответа на этот вход (третий ответ). Однако после совместной стимуляции входов I и II наблюдалось существенное увеличение величины ответа на вход II (четвертый ответ), длящееся десятки минут (рис. 12.6С). Это явление было названо ассоциативной ДВП, так как длительное увеличение ответа на вход II происходило

250 |
Раздел II. Передача информации в нервной системе |
Рис. 12.6.
Ассоциативная ДВП в срезах гиппокампа. (А) Схема эксперимента; (В) усредненные ответы; (С) суммированные результаты.
Fig. 12.6. Associative LTP in a rat hippocampal slice. (A) Intracellular records were made from a CA1 pyramidal cell while stimulating two distinct groups of presynaptic fibers in the Schaffer coUateralcommissural pathway (stimulus I and stimulus II). The stimuli were adjusted so that responses to stimulation at site I were five times greater than those to stimulation at site II. (B) Averaged responses to stimulation at site II in the control condition, after tetanic stimulation at site II (100 stimuli/s for 1 s), after a similar tetanus at site I, and after a combined tetanus at I and II. Only the combined tetanus produced potentiation; test shock 10 min later indicated a twofold increase in response amplitude. (C) Summary of the results in part B, showing the time course of the changes in response amplitude.
Stimulation at site I had no effect stimulation at II produced a brief potentiation of the response, and combined stimulation produced long-term potentiation. (After Barrionuevo and Brown, 1983.)
только при совпадении во времени стимуляции этого входа и входа I.
Механизмы индукции ДВП
Полной картины происходящих во время индукции ДПВ явлений пока еще нет. Однако все исследователи согласны с тем, что увеличение концентрации кальция в постсинаптической клетке является важным фактором. В пирамидных клетках поля СА1 индукция происходит с участием кальция, входящего через глутаматные NMDA рецепторы (глава 3). NMDA рецепторы образуют каналы для катионов с необычной характеристикой — эти каналы заблокированы при нормальном
Глава 12. Синоптическая пластичность |
251 |
потенциале покоя клетки. Канал блокирован ионами магния из экстраклеточного раствора, которые освобождают канал только при деполяризации участка мембраны с этими глутаматными рецепторами23· 24). Большая часть глутамат-чувствительных клеток экспрессирует NMDA и не-NMDA (АМРА) рецепторы в постсинаптической мембране, которые активируются при выделении глутамата из пре-синаптических терминалей25).
NMDA рецепторы обладают относительно высокой кальциевой проводимостью, однако вход кальция зависит от величины деполяризации, которая должна быть достаточна для снятия магниевого блока в каналах. Именно этим можно объяснить то, что в эксперименте по активации ассоциативной ДВП стимуляция только одного (слабого) входа II не вызывала достаточной синаптической деполяризации для снятия магниевого блока и входа кальция. Однако, когда эта стимуляция сопровождалась деполяризацией, вызванной активацией входа I, NMDA рецепторы оказались разблокированы, что привело к входу кальция и ДВП. Участие NMDA рецепторов подтверждено экспериментами, в которых антагонисты NMDA блокируют индукцию ДВП, но не мешают ДВП при применении после индукции26· 27).
Идея о том, что ДВП индуцируется увеличением концентрации кальция в постсинаптической мембране, поддержана двумя типами доказательств. Во-первых, было экспериментально показано увеличение концентрации внутриклеточного кальция во время стимуляции, при этом блокада увеличения градиента этого катиона внутриклеточно введенными буферами устраняет ДВП28· 29). Во-вторых, увеличение концентрации кальция в постсинаптической клетке благодаря извлечению его из других внутриклеточных источников приводит к длительному увеличению амплитуды ВПСП30). Таким образом, источник кальция не является существенным, поэтому в некоторых синапсах ДВП индуцируется входом кальция через потенциалзависимые кальциевые каналы, тогда как в других — высвобождением кальция из внутриклеточных депо29).
Увеличенная концентрация внутриклеточного кальция может активировать целый ряд внутриклеточных биохимических путей. Наиболее важны для индукции ДВП активация кальций/кальмодулин-зависимой киназы II (СаМКII) и цАМФ-зависимая протеинкиназы21·31). СаМКII обнаружена в больших количествах в постсинаптических уплотнениях шипиков дендритов, и внутриклеточная инъекция блокаторов СаМКII блокирует индукцию ДВП32· 33). Кроме того, индукция ДВП нарушена у мышей с генетическими нарушениями синтеза одной
из субъединиц СаМКП34). Блокада цАМФ-зависимой протеин-киназы также существенно уменьшает ДВП29· 32, 33).
Механизм проявления ДВП
Открытие ДВП поставило вопрос о том, является ли наблюдаемое увеличение амплитуды синаптического потенциала результатом увеличенного выброса передатчика из пресинаптической терминали или изменения чувствительности постсинаптической мембраны. Привлекательным казалось предположение о том, что ДВП, так же как фасилитация, усиление и ПТП, отражает увеличение квантового состава синаптического ответа, что и было показано в первых экспериментах. На рис. 12.7 показано распределение амплитуд синаптических потенциалов до и после потенциации. В ряде экспериментов статистический анализ распределений амплитуд показал увеличение среднего квантового содержания21· 35· 36). Однако в других экспериментах были показаны противоположные результаты: увеличение ВПСП происходило благодаря изменению размера одного кванта, а не количества квантов в ответе37).
Увеличение квантового содержания потенциированного ответа с одной стороны и увеличение размера кванта рассматриваются в настоящее время как указывающие на различные механизмы проявления ДВП: пресинаптический (выделяется больше квантов) и постсинаптический (больший ответ на каждый квант). Новые данные говорят в пользу того, что увеличение и квантового содержания, и числа квантов опосредованы изменениями в постсинаптической мембране.
Молчащие синапсы
Каким образом изменения в постсинаптической клетке могут проявляться в изменении количества выделяемых квантов? Было бы неправильно думать, что постсинаптическая мембрана является статической структурой, и единственным механизмом увеличения квантового содержания ПСП может быть увеличение количества квантов, выделяемых пресинаптической терминалью. Предположим, что несколько пресинаптических

252 |
|
|
|
|
|
|
|
|
Раздел II. Передача информации в нервной системе |
|
Рис. |
|
12.7. |
|
Изменения |
|
|
|
|||
квантового содержания си |
|
|
|
|||||||
наптических |
|
|
ответов |
|
|
|
||||
нейронов |
|
гиппокампа |
|
|
|
|||||
крысы при ДВП. (А, В) 16 |
|
|
|
|||||||
наложенных |
|
|
|
|
|
|
|
|||
синаптических |
токов |
до |
|
|
|
|||||
(А) |
|
и |
|
после |
(В) |
|
|
|
||
индукции ДВП. (С, D) |
|
|
|
|||||||
Распределение |
амплитуд |
|
|
|
||||||
токов до (С) и после (D) |
|
|
|
|||||||
индукции ДВП. |
|
|
|
|
|
|
||||
Fig. |
12.7. |
|
Change |
in |
|
|
|
|||
Quantum |
|
Content |
of |
|
|
|
||||
synaptic |
responses |
during |
|
|
|
|||||
LTP. Records from |
a |
rat |
|
|
|
|||||
hippocampal |
slice. |
(A, |
B) |
|
|
|
||||
Sixteen |
|
|
superimposed |
|
|
|
||||
whole-cell |
|
records |
of |
|
|
|
||||
excitatory synaptic |
currents |
|
|
|
||||||
(EPSCs) in a CA1 pyramidal |
|
|
|
|||||||
cell (A) before and (B) after |
|
|
|
|||||||
a |
conditioning |
train |
of |
|
|
|
||||
stimuli. |
Note |
the |
quan-tal |
|
|
|
||||
steps |
in |
|
the |
current |
|
|
|
|||
amplitudes. |
|
|
|
After |
|
|
|
|||
conditioning, |
the |
fraction of |
|
|
|
|||||
failures |
is |
decreased |
and |
|
|
|
||||
there |
are |
|
many |
more |
|
|
|
|||
multiquantal |
responses. (C, |
|
|
|
||||||
D) |
Distribution |
of |
current |
|
|
|
||||
amplitudes |
(C) |
before |
and |
|
|
|
||||
after |
(D) conditioning. |
The |
|
|
|
|||||
normal curve is fitted to |
|
|
|
|||||||
baseline |
noise |
(failures); |
|
|
|
|||||
|
|
|||||||||
arrows |
indicate |
mean |
|
produced by a single quantum. After potentiation, the number of failures is reduced and |
|
|||||
current |
|
|
|
|
|
|
|
the mean current is increased in amplitude by a factor of almost three, while the single- |
|
|
|
|
|
|
|
|
|
|
|
quantum current is unchanged. (After Malinow and Tsien, 1990.) |
|
бутонов контактируют с постсинаптической мембраной дендритного шипика который содержит всего несколько работающих АМРА рецепторов, или их совсем нет. В условиях покоя выброс глутамата в таком синапсе вызовет очень маленький или никакого ответа
— такой синапс будет «молчащим»21). Предположим, после индукции ДВП новые глутаматные АМРА рецепторы будут включены в постсинаптическую мембрану молчащего синапса. Эти рецепторы начнут отвечать на кванты медиатора, выбрасываемые из пресинаптической терминали, и квантовый состав ответа соответственно увеличится (рис. 12.8).
Регуляция количества синаптических рецепторов
В настоящее время существуют доказательства того, что при вызове ДВП АМРА рецепторы увеличиваются в количестве. Показано, что субъединицы АМРА рецепторов доставляются в шипики дендритов после ритмической стимуляции, сопровождающейся активацией NMDA рецепторов38). Субъединица АМРА рецептора GluRl (см. главу 3), меченая зеленым флуоресцентным пептидом (GFP), была экспрессирована в нейронах гиппокампа в культуре. Анализ распределения меченой субъединицы в отростках с помощью лазерной сканирующей микроскопии и электронной микроскопии показал, что большая часть GluRl-GFP находится в дендритах. После стимуляции нейронов, меченые рецепторы быстро появились и в тех дендритах, где их раньше не было. Эти данные позволяют предположить, что многие дендритные шипики являются молчащими и получают полный набор АМРА рецепторов после ритмической стимуляции.
Дополнительные свидетельства увеличения количества АМРА рецепторов были сум-