

Nikolls_-_Ot_neyrona_k_mozgu
.pdfГлава 18. Обработка соматосенсорных и слуховых сигналов |
393 |
Рис. 18.6. Соматосенсорная кора. Подотдел первичной соматосенсорной коры (S1) показан на сагиттальном срезе, проходящем через постцентральную извилину коры мозга обезьяны. S, подразделяется на зоны 3а, Зb, 1 и 2, лежащие позади центральной борозды. Зона 5 является частью соматосенсорной ассоциативной коры. Зона 4, лежащая спереди от центральной борозды, является первичной моторной корой.
Fig. 18.6. Somatosensory Cortex. The subdivisions of primary somatosensory cortex (5г) are shown here on a sagittal section through the postcentral gyrus of the monkey cortex. S is subdivided into areas За, ЗЪ. 1, and 2. proceeding posteriorly from the central sulcus. Area 5 is part of the somatosensory association cortex. Area 4 lying anterior to the central sulcus is primary motor cortex.
личения границ. Стимул, который приходится как на возбужденный центр, так и на тормозное окружение, будет менее эффективен, чем стимул, который прекращается (имеет границу) в пределах возбужденного центра. Это, опять же, иллюстрирует тот факт, что нервная система опирается на распознавание изменений, в данном случае — пространственных границ.
Параллельная обработка сенсорных модальностей
Передне-центральная извилина может быть разделена на несколько областей на основе различий в клеточной организации и сенсорной модальности (рис. 18.6). Область За является непосредственно смежной с первичной моторной корой, и нейроны там реагируют прежде всего на мышечные рецепторы растяжения и на другие глубокие рецепторы 17) В противоположность этому, область 3b принимает входы от быстро и медленно адаптирующихся кожных рецепторов, особенно тех, которые имеют малые рецептивные поля. Специфическая локализация рецепторного типа в пределах подобластей S1 иллюстрирует концепцию параллельной обработки в каждой сенсорной системе. Каждая область тела представлена в кожной зоне (3b), так же как и в зоне глубоких рецепторов (3a). Таким образом, соматотопическая корковая карта фактически состоит из множества представительств плана тела, по одному для каждой сенсорной модальности.
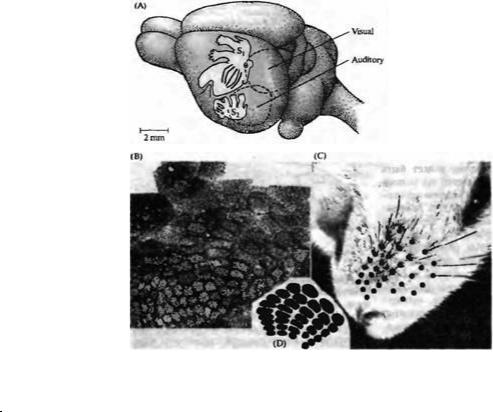
Точная анатомическая и функциональная организация соматосенсорной коры особенно ярко видна на примере проекции сенсорной иннервации от вибрисс мыши (рис. 18.7). Как и у людей, лицевая область коры у мыши непропорционально велика. Гистологические исследования Вулси и Ван дер Луза18) показали, что эта область соматосенсорной коры у мыши содержит характерные группы нервных клеток, собранных вместе в пучки в форме цилиндров, которые лежат перпендикулярно в коре. Эти скопления были названы бочонками за свою форму, которая была выявлена путем реконструкции по серийным срезам. Каждый бочонок — от 100 до 400 мкм в диаметре — состоит из кольца клеток, окружающих сердцевину, содержащую немногочисленные клетки.
Массив бочонков у мыши обычно организован в пять рядов. Замечательно, что этот паттерн точно соответствует рядам вибрисс на морде мыши — один бочонок для одного уса. Если ус удалить сразу после рождения, соответствующий бочонок исчезает из коры19). Деафферентация на более поздних этапах приводит к разнообразным изменениям, включая измененную экспрессию рецепторов нейромедиаторов в «бочоночной» коре20· 21). Функциональное значение бочонков подтверждено электрическим отведением от индивидуальных клеток коры22· 23). Каждая клетка отвечает на движение соответствующего уса — некоторые избирательно возбуждаются при движении уса в конкретном направлении. Таким образом, значительная часть соматосенсорной коры мыши занята обработкой информации, поступающей от механорецепторов, расположенных на морде. Мышь использует свои вибриссы как чувствительные антенны, двигая ими вперед и назад,
394 |
|
|
|
|
|
Раздел III. Интегративные механизмы |
Рис. 18.7. Бочонки в |
|
|||||
соматосенсорной |
|
коре |
|
|||
мыши, |
соответствующие |
|
||||
вибриссам. (А) Схема мозга |
|
|||||
мыши, |
показывающая |
|
||||
соматотопическое |
|
|
|
|||
представительство |
|
в |
|
|||
сенсорной |
|
|
|
коре |
|
|
(«мускулус», |
|
|
который |
|
||
соответствует |
|
гомункулусу |
|
|||
на рис. 18.3В), в частности, |
|
|||||
морды и вибрисс. (В) |
|
|||||
Горизонтальной |
срез |
коры |
|
|||
мозга |
через |
|
область, |
|
||
представляющую вибриссы, |
|
|||||
показывающий |
бочонки в |
|
||||
поперечном сечении. (С) |
|
|||||
Увеличенный |
вид |
вибрисс, |
|
|||
место |
|
возникновения |
|
|||
каждой |
из |
них |
показано |
|
||
черной |
точкой. |
(D) |
|
|||
Схематическое |
|
|
|
|
||
изображение |
расположения |
|
||||
бочонков |
|
|
на |
|
В, |
|
показывающее |
|
|
их |
|
||
соответствие вибриссам. |
|
|||||
Fig. 18.7. Barrels in Mouse |
|
|||||
Somatosensory |
|
|
Cortex |
|
||
corresponding |
to |
vibrissae. |
|
|||
(A) Diagram of a mouse brain |
|
|||||
showing |
the |
|
somatotopic |
|
||
representation on the sensory |
|
|||||
cortex (the "musculus," which |
S2 is the secondary somatosen sory cortex. (B) Horizontal section through the cortex in |
|||||
corresponds |
|
to |
|
the |
||
homunculus in Figure 18.3B), |
the area representing the vibrissae, showing barrels in cross section. (C) Close-up of |
|||||
particularly the |
face |
and |
whiskers, the origin of each marked by a black dot. (D) Schematic diagram of barrel |
|||
vibrissae. |
|
|
|
|
|
arrangement from B, showing correspondence with vibrissae. (After Woolsey and Van |
|
|
|
|
|
der Loos, 1970.) |
|
|
|
|
|
|
|
|
по мере того, как она движется, чтобы обнаруживать объекты по обе стороны своего пути24).
Вторичная и ассоциативная соматосенсорная кора
Множественное представительство тела находится в первичной соматосенсорной коре (S1). По мере удаление от центральной борозды, от области 3b до 1 и до 2, рецептивные поля становятся больше, и организация по принципу «центр — окружение», избирательность по направлению и интеграция субмодальности становятся все более выраженными в нейронном ответе25, 26) . Область 1 лежит непосредственно позади области 3b и получает от нее активирующий вход. Область 2 получает вход как от 3a, так и от 3b.
В дополнение к первичной соматосенсорной коре, множественные представительства тела имеются в других, вторичных областях. В общем это зоны или полосы коры, которые лежат позади и вентролатеральнее по отношению к S1. У приматов вторичная соматосенсорная кора (S2) лежит на передней стенке боковой борозды, находясь вентральнее, чем S1 (рис. 18.8). S2 также соматотопически организована, и получает вход как от таламуса, так и от S1. Если у приматов хирургически удалить S1, активность клеток в S2 значительно снижается27). Если из S1 убраны только области 3b и 2 (где обрабатывается поверхностная, или кожная, стимуляция), то количество клеток в S2, отвечающих на кожную стимуляцию, специфически уменьшается28). Результаты этих экспериментов свидетельствуют, что соматосенсорная информация поступает от S1 к S2. У других видов прямой таламический путь к S2 имеет более важное значение29).
Поток информации от S1 к S2 также может быть иерархическим в том смысле, что эта информация обрабатывается в S1, a S2 затем имеет дело уже с производными свойствами соматочувствительности. Например, когда обезьяна прикасается кончиками пальцев к вращающемуся барабану, во многих нейронах в S1 импульсация возрастает пропор-

Глава 18. Обработка соматосенсорных и слуховых сигналов |
395 |
Рис 18.8. Соматосенсорные области коры. Первичная соматосенсорная кора S лежит в постцентральной извилине непосредственно позади центральной борозды. Соматосенсорные ассоциативные области
коры (зоны 5 и 7 по Бродману) лежат позади от S1. Вторичная соматосенсорная кора S2 лежит ниже S1, в складке боковой борозды.
Fig. 18.8. Somatosensory Cortices. The primary
somatosensory cortex, S1, lies on the postcentral gyms immediately behind the central sulcus Somatosensory association cortices (Brodmann's areas 5 and 7) lie
posterior to S1. The secondary Somatosensory cortex, S2, lies below S1, tucked into the lateral sulcus.
ционально увеличению неровности поверхности барабана, как если бы они управлялись непосредственно афферентным входом. В S2, однако, многие нейроны сигнализируют об изменении текстуры, независимо от величины этого изменения30). Другими словами, нейроны в S2 выводят концепцию (различие в качестве поверхности) из более простой информации, поступающей в S1. Другие наблюдения также свидетельствуют, что зона S2 имеет дело со сложными аспектами сенсорного кодирования. Например, множество клеток в S2 y приматов имеют билатеральные рецептивные поля31), отвечая на стимуляцию сходных областей на обеих сторонах тела.
Области 5 и 7 по Бродману представляют собой соматосенсорную ассоциативную кору. Нейроны, расположенные здесь, реагируют на стимуляцию кожи или движение конечности, но требования к стимулу у них сложны и отличаются от профилей реакции первичных сенсорных афферентов. Более того, нейроны здесь могут также реагировать на зрительную стимуляцию. Хотя специфические функции вспомогательных областей соматосенсорной коры до конца не понятны, ясно то, что тенденция к более сложно организованным стимулам в них возрастает по мере удаления от центральной борозды.
Болевые и температурные проводящие пути
Информация о вредоносных стимулах и температуре передается в высшие центры специфическими рецепторами и по путям, в значительной степени отличным от ощущений положения, прикосновения и давления. Ощущения боли и температуры передаются по миелинизированным и немиелинизированным афферентам малого диаметра, имеющим свободные нервные окончания в коже и других тканях. Окончания морфологически не специализированы, но могут быть разделены на разные подтипы, лучше реагирующие на осязательные стимулы, нагрев или охлаждение, повреждение и боль32). Ноцицептивные и температурно-чувствительные афферентные аксоны подразделяются по величине на два класса. А-δ-волокна имеют от 1 до 4 мкм в диаметре и проводят возбуждение со скоростью от 6 до 25 м/с, а немиелинизированные С-аксоны от 0,1 до 1 мкм в диаметре — со скоростью от
0,5 до 2 м/с (глава 7).
Два типа волокон передают различные болевые ощущения. У человека короткий, интенсивный стимул, подаваемый на удаленный участок конечности, вызывает сначала острое и относительно короткое колющее болевое ощущение (первичная боль), за которым следует тупое, длительное ощущение жжения (вторичная боль). Электрофизиологические эксперименты показали, что первичная боль связана с активацией мелких миелинизированных волокон, а вторичная боль — с активацией С-волокон33)--36).
Центральные пути боли
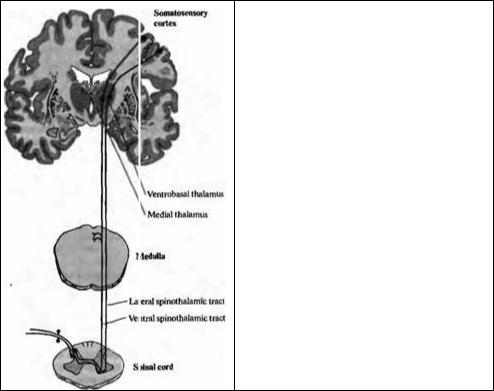
Ноцицептивные и температурно-чувствительные афферентные аксоны образуют синапсы на клетках второго порядка внутри задних столбов спинного мозга. Волокна от клеток второго порядка переходят на противоположную сторону и восходят по двум основным путям: латеральному и вентральному спиноталамическим трактам (рис. 18.9). Восходящие волокна оканчиваются в вентробазальном и медиальном ядрах таламуса — поэтому они называются спиноталамическим трактом. Клетки в вентробазальном и медиальном ядрах таламуса проецируются на соматосенсорную кору и далее по всему мозгу. В отличие от нейронов высокого порядка в задних столбах (лемнисках), пути в таламу-се и коре, получающие Ноцицептивные вхо-
396 |
Раздел III. Интегративные механизмы |
Рис 18.9. Сенсорные проводящие пути, передающие болевые и температурные ощущения. Мелкие миелинизированные и немиелинизированные афферентные волокна оканчиваются ипсилатерально на сегментарных спинномозговых нейронах, аксоны которых идут контралатерально в составе вентрального и латерального спиноталамических трактов, оканчиваясь в медиальных и вентробазальных ядрах таламуса Нейроны третьего порядка затем проецируются в кору мозга. Волокна вентрального спиноталамического тракта дают коллатеральные ответвления в продолговатом мозге, а некоторые из них оканчиваются на зтом уровне (спи норетикулярный тракт не показан).
Fig. 18.9. Sensory Pathways Mediating Pain and Temperature, shown on cross sections of the spinal cord and medulla and a coronal section of the brain. Small myelinated and unmyelinated fibers enter the spinal cord to form synapses with second-order neurons. The second-order axons cross the midline to ascend in the ventral and lateral spinothalamic tracts to end in the medial and ventrobasal nuclei of the thalamus. Third-order cells then project to the cerebral cortex. Cells in the ventral spinothalamic tract give off collateral branches in the medulla, and some terminate at this level (the spinoreticular tract; not shown).
ды, обладают большими, нечетко локализованными рецептивными полями, часто билатерально покрывающими обширные области тела38).
Хотя афферентные ноцицепторы имеют свои собственные пути в коре, ясно, что вредоносные стимулы могут распространяться по тем же восходящим путям в составе задних столбов (лемнисков), которые проводят возбуждение от рецепторов других модальностей. Действительно, многочисленные эксперименты показали, что две соматосенсорные системы взаимодействуют. В частности, легкое прикосновение или постукивание по коже может повлиять на разряд ноцицептивных нейронов в центральной нервной системе. Активация (постукиванием) соматосенсорных афферентов, имеющих перекрывающиеся рецептивные поля, вызывает тормозное влияние на ноцицептивные разряды коревых или таламических нейронов39), что позволяет предположить, что синаптическое взаимодействие в спинном мозге «открывает ворота» болевым ощущениям40).
Синаптическая передача между болевыми афферентами и спиноталамическими нейронами второго порядка в составе заднего рога тормозится интернейронами, которые действуют как пре-, так и постсинаптически (рис. 18.10). Эти тормозные интернейроны могут быть активированы коллатералями афферентов большого диаметра (лемнисковые афференты). Таким образом, рецепторы осязания в коже могут подавить ощущение боли путем такого взаимодействия. Тема модуляции болевого ощущения привлекла к себе значительное внимание с открытием опиоидных рецепторов в центральной нервной системе и с определением природных опиато-подобных пептидов в специфических нейронах, выполняющих ноцицептивную функцию1) (см. обсуждение пептидных нейротрансмиттеров в главе 14)41).
Как в других сенсорных системах, нисходящие влияния от высших центров могут серьезно изменить поток сенсорной информации о вредоносных стимулах, который достигает сознания42· 43). Обсуждение конкретных путей, обеспечивающих эти влияния,

Глава 18. Обработка соматосенсорных и слуховых сигналов |
397 |
|||
Рис. 18.10. Соматосенсорное |
торможение |
|
||
передачи |
боли. |
Синаптическая |
|
|
передача от болевых афферентов на |
|
|||
нейроны второго порядка в спинном мозге |
|
|||
уменьшается |
|
|
тормозными |
|
интернейронами. |
|
Тормозные |
|
|
интернейроны |
могут |
активироваться |
|
|
сенсорными |
афферентными |
волокнами |
|
|
большого диаметра (которые передают |
|
|||
ощущения тонкого прикосновения или |
|
|||
вибрации). |
|
|
|
|
Fig. 18.10. Somatosensory Inhibition of Pain Transmission. Synaptic transmission from pain afférents to second-order neurons in the spinal cord is reduced by inhibitory interneurons. The inhibitory interneurons can be activated by collaterals of large-diameter sensory fibers (such as those that mediate fine touch or vibration sense).
можно найти в содержательной монографии Филдза44) о нейрофизиологических и клинических аспектах боли. Этот путь начинается в среднем мозге от группы клеток серого вещества периаквелуктальной области. Как было обнаружено в некоторых случаях, стимуляция этой области имплантированными электродами вызывает избирательное снижение сильной клинической боли45). Считается, что задействованные в этом клетки являются энкефалинергическими и проецируются на серотонинергические нейроны в ростральной части ствола мозга (rostral medulla). Стволовые нейроны затем посылают нисходящие волокна по дорсолатеральному фуникулярному тракту спинного мозга в задние рога, а именно в I и II слои. Там они образуют синапсы с интернейронами и терминалями афферентных волокон, модулируя передачу в болевых путях. Нисходящие волокна сопровождаются норадренергическими волокнами, принадлежащими ко второму пути, берущему начало в дорсолатеральном отделе моста, который также задействован в модуляции боли.
Сама боль остается нечетким и сложным понятием, выходящим за пределы данной книги. В отличие от анализа зрительной, слуховой и соматосенсорной систем, обсуждение боли со всем ее высоким эмоциональным содержанием неизбежно сталкивается с рассмотрением субъективных вопросов, чувств, подобных «горю» и «страданию», которые в настоящее время не могут быть выражены языком нейробиологии (точно так же, как «созерцание заката» или «чувство тепла» не может рассматриваться в этих терминах). Тем не менее определенные соотношения между нервной деятельностью и болью очевидны.
Так, общеизвестное переживание острой, четко локализованной приступообразной боли, после которой следует тупое, постепенно растекающееся, плохо локализованное жжение, может быть приписано активности волокон А-δ и С. Подобным же образом обезболивающий эффект постукивания по коже может быть объяснен синаптическими взаимодействиями на уровне спинного мозга, таламуса и коры. Особенно притягательна идея, что уровень воспринимаемой боли может быть снижен нисходящими влияниями, которые включают в себя действие морфиноподобных пептидов, дающих возможность солдату, который бросается в битву, не замечать ран и повреждений, которые при нормальных обстоятельствах были бы чрезвычайно болезненны.
§ 2. Слуховая система: кодирование частоты звука
Соматосенсорная система анализирует стимулы на основе их расположения на карте тела. Благодаря аналогичному процессу, зрительные стимулы являются ретинотопически картированными и анализируются на предмет их местоположения в визуальном (пространственном) мире. Однако слуховая система использует совершенно иной метод анализа. Слуховая периферия, или улитка, картирует звуковые стимулы скорее по их частотному содержанию, чем по пространственному расположению, и последующие стадии слуховой системы используют этот тонотопический анализ, чтобы определить смысл речи и значение других сложных звуков. Как частота звука кодируется волосковыми клетками улитки, и как эти сигналы обрабатываются
398 Раздел III. Интегративные механизмы
на более высоких уровнях слуховой системы? Как определяется пространственное расположение звука?
У наземных позвоночных звуковые волны в воздухе входят во внешнее ухо, колеблют барабанную перепонку и, благодаря серии механических сопряжений в среднем ухе, конвертируются в колебания жидкости улитки. Колебания жидкости, в свою очередь, вызывают вибрацию базилярной мембраны, на которой находятся сенсорные волосковые клетки в составе органа Корти. Этот процесс описан поэтически Олдосом Хаксли:
Трубный звук Понгилеони и скрежет безымянных скрипок потрясли воздух в большой зале, вызвали дрожь в оконных стеклах, и это, в свою очередь, сотрясло воздух в апартаментах лорда Эдварда на дальней стороне. Сотрясающийся воздух задребезжал в барабанных перепонках лорда Эдварда; соединенные между собой косточки молоточка (ушной косточки), наковальни внутреннего уха и стремени (слуховой кости среднего уха) были приведены в движение, так что возбудили мембрану окна (oval window) и подняли бесконечно малую бурю в жидкости лабиринта. Волосковые окончания слухового нерва дрожали, как водоросли в бушующем море; огромное количество непонятных чудес произошло в мозгу, и лорд Эдвард в экстазе прошептал: «Бах!»
Улитка
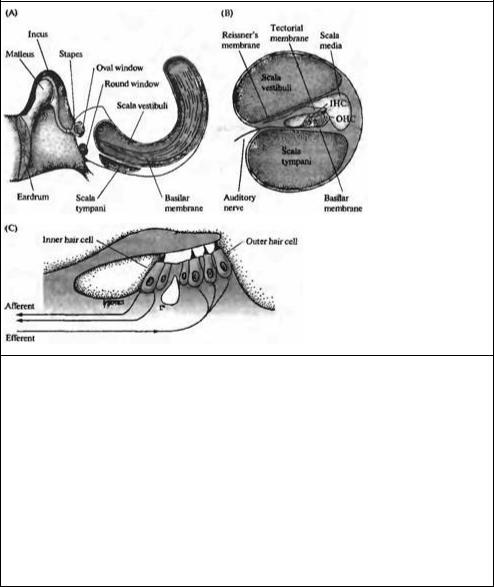
Канал улитки разделен на три отдела. Средняя лестница (scala media) содержит раствор с высоким содержанием калия, эндолимфу. Он отделен от покрывающей его вестибулярной лестницы (scala vestibuli) Рейсснеровой мембраной, а от наполненного жидкостью пространства барабанной лестницы (scala timpani) — тесными межклеточными соединениями между апикальными окончаниями волосковых клеток и окружающими вспомогательными клетками (рис. 18.11В). Барабанная и вестибулярная лестницы содержат перилимфу, сходную по составу с цереброспинальной жидкостью; средняя лестница содержит эндолимфу, ионный состав которой похож на состав цитоплазмы: высокое содержание калия, низкое — натрия и с концентрацией кальция, поддерживаемой на микромолярном уровне47· 48). Необычный состав внеклеточной жидкости устанавливается благодаря работе ионных насосов в клетках, расположенных в сосудистом слое (stria vascularis), т. е. в секреторном эпителии, выстилающем боковую стенку средней лестницы.
Существуют две различные группы волосковых клеток в улитке млекопитающих: внутренние волосковые клетки и внешние волосковые клетки. Они различаются по своему расположению (внутренние волосковые клетки ближе к центральной оси спирали улитки) и по паттерну иннервации. Внутренние волосковые клетки получают более 90% афферентных контактов улитки49)--51); внешние волосковые клетки являются постсинаптическими мишенями для эфферентной иннервации (рис. 18.11С)52). Эти различия в паттернах иннервации ставят интересные вопросы, касающиеся функциональной роли этих двух типов клеток, которые будут обсуждаться далее в этой главе. Покровная мембрана (tectorial membrane) налегает на волосковые пучки как внутренних, так и внешних волосковых клеток, и различие в движении покровной и базилярной мембран приводит к боковому сдвигу, открывающему каналы механотрансдукции (как описано в главе 17).
Частотная избирательность: механическая настройка
Звуковое восприятие — это не просто вопрос выявления звукового стимула. Информационное содержание звука является функцией его частотного состава. Таким образом, слуховая система зависит от способности ее механосенсорных волосковых клеток быть частотноизбирательными. В улитке млекопитающих настройка по частоте зависит от механических свойств специализированной структуры, в которой находятся сенсорные клетки, аналогично механической адаптации, осуществляемой многослойной капсулой в тельце Пачини. Ширина и толщина базилярной мембраны закономерно меняется по длине канала улитки (рис. 18.11 А). У основания улитки (ближе к овальному окну) базилярная мембрана узкая и жесткая; у противоположного конца (вершина улитки) она широкая и гибкая. По причине этой изменяющейся жесткости входящие звуковые волны вызывают движение жидкости и вибрацию базилярной мембраны в разных участках в зависимости от частоты звука.
Это свойство функции улитки было описано фон Бекеси53), который использовал стробоскопическое освещение отражающих частиц, рассеянных в улитке, чтобы визуализировать паттерн вибрации. Он обнаружил, что звуки высокой частоты вызывают максимальные колебания на более толстой, жест-
Глава 18. Обработка соматосенсорных и слуховых сигналов |
399 |
|
|
Рис. 18.11. Структура улитки. (А) Схема среднего уха и улитки, показывающая систему косточек, с помощью которых барабанная перепонка соединена с мембраной овального окна улитки. Улитка дана рассеченной и не скрученной, чтобы показать ее основные внутренние полости (вестибулярную и барабанную лестницы) и форму базилярной мембраны, расширяющейся от основания улитки к ее вершине. (В) Поперечный разрез улитки, показывающий структурные взаимоотношения между базилярной мембраной, внутренними (IHC) и наружными (ОНС) волосковыми клетками и покровной мембраной. (С) Иннервация волосковых клеток улитки. Около 95 % афферентных волокон являются постсинаптическими мишенями для IHC. До 20 афферентных волокон контактирует с одиночной IHC. ОНС имеют мало афферентных связей, зато являются постсинаптическими мишенями для холинергических эфферентных нейронов, которые проецируются из оливарного комплекса, расположенного в стволе мозга.
Fig. 18.11. Structure of the Cochlea. (A) The middle ear and cochlea, showing the eardrum and its bony connections to the oval window. The cochlea is represented as unwound and cut open to show the major internal compartments (scala vestibuli and scala tympani), and overlying structures have been removed to reveal the shape of the basilar membrane, which is narrow near the cochlear base and broad at the apex. (B) Cross section of the cochlea, showing the scala media (which contains endolymph, a high-potassium solution) and the structural relations among the basilar membrane, inner hair cells (IHC) and outer (OHC) hair cells, and tectorial membrane. Hair cells form synapses on the terminals of auditory nerve fibers that have their cell bodies in the spiral ganglion. (C) Innervation of cochlear hair cells. Approximately 95 % of afferent fibers are po tsynaptic to IHCs. As many as 20 afferent fibers contact a single IHC. OHCs have few afferent contacts but instead are the postsynaptic targets of cholinergic efferent neurons that project from the olivary complex in the brainstem.
кой нижней части мембраны, а звуки низкой частоты вызывают максимальную вибрацию на более гибкой верхней части мембраны улитки. Фон Бекеси назвал этот паттерн колебаний «бегущей волной».
Как следствие механической настройки базилярной мембраны, волосковые клетки и их постсинаптические афферентные волокна у основания улитки преимущественно стимулируются высокочастотным звуком, тогда как те, что находятся у вершины улитки, лучше всего реагируют на низкочастотный звук (рис. 18.12). Частотная избирательность афферентов улитки может быть измерена при помощи отведения потенциалов действия, возникающих в ответ на чистые тона звука,

400 |
|
|
|
Раздел III. Интегративные механизмы |
|
|||
Рис. 18.12. Настройка улитки. (А) Локализация |
|
|
||||||
участков улитки, в которых звуковые волны |
|
|
||||||
вызывают максимальное смещение базилярной |
|
|
||||||
мембраны, зависит от частоты. Кривые |
|
|
||||||
представляют |
относительное |
смещение на |
|
|
||||
указанных частотах (100-2000 Гц). (В) Типичные |
|
|
||||||
кривые настройки |
четырех |
индивидуальных |
|
|
||||
волокон слухового нерва, иннервирующих |
|
|
||||||
различные |
участки |
улитки. |
Показана |
|
|
|||
зависимость между частотой звукового стимула |
|
|
||||||
и интенсивностью звука в децибелах (dB) |
|
|
||||||
необходимой для возникновения разряда в |
|
|
||||||
волокне. Пики соответствуют оптимальным |
|
|
||||||
звуковым частотам (т. е. частотам, при которых |
|
|
||||||
интенсивность стимула для каждого волокна |
|
|
||||||
минимальна). |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
Fig. 18.12. Cochlea |
Tuning. (A) The location of |
|
|
|||||
maximum displacement of the basilar membrane in |
|
|
||||||
the cochlea by sound waves depends on |
|
|
||||||
frequency. The |
curves |
represent |
relative |
|
|
|||
displacement at the indicated frequencies (100-2 |
|
|
||||||
000 Hz). At the low frequencies, maximum |
|
|
||||||
displacement is near the wider (more flexible) |
|
|
||||||
apical membrane; higher frequencies produce |
|
|
||||||
maximal displacement near the narrower (stiffer) |
|
|
||||||
base. (B) Typical tuning curves of four individual |
|
|
||||||
eighthnerve fibers innervating different locations on |
|
|
||||||
the cochlea. Sound intensity in decibels (dB) |
|
|
||||||
The best frequencies for the fibers (i. e., |
the frequencies |
|||||||
needed to produce discharges in a fiber is plotted |
||||||||
requiring the least stimulus intensity) are 1 000, 2 000, 5 000, |
||||||||
against frequency of the auditory stimulus. |
|
|||||||
|
and 10 000 Hz. (A after von Bekesy, 1960; |
В after Katsuki, |
||||||
|
|
|
|
|
|
|||
|
|
|
|
|
|
1961.) |
|
|
частота и интенсивность которых систематически варьирует. Результирующая кривая настройки V-образна, с оптимальной, или характеристической, частотой, определяемой как чистый тон, к которому волокно наиболее чувствительно. Характеристическая частота каждого волокна определяется по тому, где именно в канале улитки оно контактирует с волосковой клеткой.
Звуковая частота картируется на протяжении эпителиального массива рецепторов, производящих кохлеотопическую, или тонотопическую карту частоты. Поэтому, слуховая система может использовать принципы нейронной обработки — такие, как латеральное торможение — основанные на тонотопии, чтобы истолковывать и обрабатывать информацию, закодированную в звуковой частоте таким же образом, как соматотопические карты лежат в основе обработки соматосенсорных сигналов.
Эфферентное торможение улитки
Как и мышечные веретена, слуховые рецепторы подвергаются эфферентной регуляции.
Нейроны комплекса верхних олив (olivary complex) в стволе мозга млекопитающих проецируются на ипсилатералъную и контралатеральную улитку54'. Активация этого пути вызывает высвобождение ацетилхолина в эфферентных синапсах на волосковых клетках и подавляет ответы на звук в афферентных волокнах улитки55· 56). Эфферентная обратная связь снижает чувствительность улитки при наличии фонового шума и снижает риск перегрузки57). Это — аналог переустановки чувствительности мышечного веретена гамма-эфферентами, поддерживающими поток информации во время движения конечности в широком диапазоне положений (глава 22). Эфферентные волокна активируются звуком и иннервируют ограниченные области улитки; в результате подавление шума носит частотноспецифический характер58)--61). В дополнение к восстановлению динамического диапазона торможение расширяет кривую настройки афферентных волокон. Наконец, эфферентная обратная связь может защитить улитку от повреждения громким звуком62). Фактически, сила эфферентной обратной связи находится в обратном соотношении со степенью
Глава 18. Обработка соматосенсорных и слуховых сигналов |
401 |
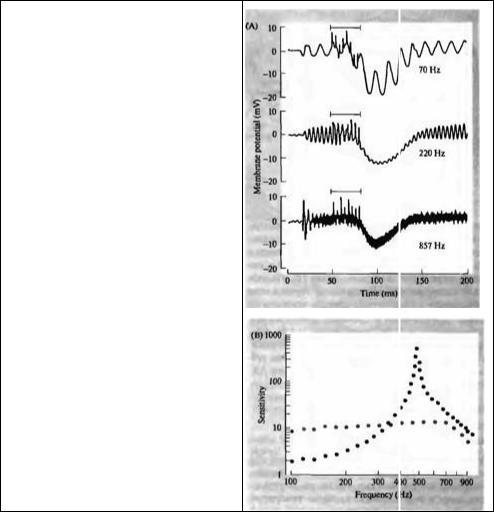
Рис. 18.13. Влияние эфферентной стимуляции волосковых клеток улитки на звуковые стимулы. (А) Средняя запись: осцилляторный ответ волосковой клетки на звуковой стимул частотой 220 Гц (резонансная частота) тормозится короткой пачкой эфферентных стимулов (обозначено полоской), и клетка гиперполяризуется. Верхняя запись: ответ на звуковой стимул при 70 Гц. Интенсивность стимула подобрана таким образом, что осцилляторный ответ был близок по амплитуде ответу на звуковой стимул частотой 220 Гц. Эфферентная стимуляция также вызывает гиперполяризацию, однако не ослабляет, а, наоборот, усиливает осцилляторный ответ. Нижняя запись: то же при частоте 857 Гц, однако этот ответ не изменяется при эфферентной стимуляции. (В) Чувствительность другой волосковой клетки (в мВ на единицу звукового давления) показана как функция частоты при эфферентной стимуляции (серым цветом) и в ее отсутствии (черным). Эфферентное торможение снижает ответ при резонансной частоте и повышает чувствительность на более низких частотах, в результате чего настройка нарушается.
Fig. 18.13. Effect of Efferent Stimulation on cochlear hair cell responses to acoustic stimuli. (A) Center record: The oscillatory response of a cell to an acoustic stimulus of 220 Hz (its resonant frequency) is inhibited by a brief train of efferent stimuli (indicated by the bar), and the cell is hyperpolarized. Upper record: The response to an acoustic stimulus at 70 Hz. The stimulus intensity was adjusted so that the oscillatory response was similar in magnitude to that at 220 Hz. Efferent stimulation produces a hyperpolarization, but an increase rather than a decrease in the oscillatory response. Lower record: The oscillatory response to an 857 Hz stimulus (again adjusted to produce a response similar to that at the resonant frequency) is unchanged by efferent stimulation. (B) Sensitivity of another cell (in millivolts per unit of sound pressure) as a function of frequency in the absence (grey) and presence (black) of efferent stimulation. Efferent inhibition reduces the response at the resonant frequency and increases the sensitivity at low frequencies, thereby degrading tuning. (A after Art et al., 1985; В after Fettiplace, 1987.)
акустического повреждения, вызванного громким звуком63).
Активация эфферентного пути к уху черепахи вызывает большие гиперполяризирующие тормозные постсинаптические потенциалы (IPSP) в волосковых клетках64· 65). Влияние торможения на ответ волосковой клетки на звук показан на рис. 18.3А. Клетка была стимулирована чистыми тонами на трех частотах, одна из которых является оптимальной для данной болосковой клетки, вторая — более высокой частотой, и третья — более низкой частотой. Интенсивности тонов были отрегулированы так, что все они вызывали осциллирующий рецепторный потенциал одной и той же амплитуды. Короткая серия стимулов, поданная на эфферентное волокно, гиперполяризировала клетку и сильно ослабила ее ответ на тон при 220 Гц (характеристическая частота). На более низкой и высокой частотах акустической стимуляции активация эфферентов по-прежнему приводила к гиперполяризации, но низкочастотная осцилляция даже увеличивалась по амплитуде, тогда как на высоких частотах амплитуда была неизменна. Этот дифференциальный эффект торможения приводит к расширению частотного диапазона ответов волосковых клеток (рис. 18.13В).
Механизм холинергического торможения был исследован в волосковых клетках цыпленка66, 67). Ацетилхолиновые (АХ) рецепторы
402 |
Раздел III. Интегративные механизмы |
Рис. 18.14. Электрическая подвижность наружных вол сковых клеток. Для фиксации мембранного потенциала наружных волосковых клеток, изолированных из улитки позвоночных, были использованы пэтч пипетки Деполяризация приводила к укорочению клеток; гилерполяризация делала их длиннее. Такие изменения длины могли быть достаточно веяики, достигая 30 нм/мВ.
Fig. 18.14. Electromotility of Outer Hair Cells. Whole-cell patch pipettes are used to clamp the membrane potential of outer hair cells isolated from the mammalian cochlea. Depolarization causes the cells to shorten; hyperpolarization makes them longer. Such length changes can be as large as 30 nm/mV.
являются лиганд-регулируемыми катионными каналами, через которые натрий и кальций входят в клетку, приводя к активации кальцийзависимых калиевых каналов и к гиперполяризации мембраны (глава 10). Подобный холинергический механизм торможения обнаружен в волосковых клетках млекопитающих68)--70). Ответы волосковых клеток на АХ блокируются α-бунгаротоксином, и есть надежное доказательство того, что необычный член семейства никотиновых рецепторов а9 является лиганд-связующей субъединицей АХ рецептора волосковой клетки71)--73).
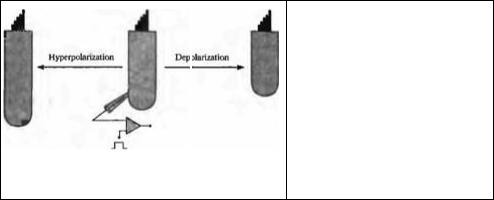
Электрическая подвижность волосковых клеток улитки млекопитающих
Эфферентные нейроны, которые тормозят ответы улитки, образуют синапсы на внешних волосковых клетках, но не на внутренних волосковых клетках, которые генерируют нейрональный выход улитки млекопитающих. Как этот тормозящий эффект передается от внешней к внутренней волосковой клетке? Важное свойство внешней волосковой клетки в улитке млекопитающих — электрическая подвижность. Внешние волосковые клетки, изолированные из улитки млекопитающих, укорачиваются, реагируя на деполяризацию, и удлиняются во время гиперполяризации их мембран (рис. 18.14)74· 75). Эти движения не вызываются актин-миозином, но возникают в результате прямых эффектов электрического потенциала на заряженные «моторные» протеины в базолатеральной мембране76)--78). Каков бы ни был механизм электрической подвижности внешних волосковых клеток, она также способствует вибрации структуры улитки во время звуковой стимуляции, и, таким образом, увеличивает отклонение стереоцилиев внутренних волосковых клеток79).
Внешние волосковые клетки улучшают настройку улитки, сообщая дополнительную механическую энергию движению базилярной мембраны (рис. 18.15). Это объясняет, как эфферентные синапсы, образованные на внешних волосковых клетках, подавляют звуковой ответ внутренних волосковых клеток. Поскольку подвижность внешних волосковых клеток вызывается изменениями мембранного потенциала, синаптическое торможение действует так, что подавляет электрическую подвижность аналогично тормозным эффектам в других областях нервной системы. Торможение уменьшает активный механический вклад внешних клеток в колебания структур улитки и, таким образом, опосредованно снижает возбуждение внутренних волосковых клеток.
Электрическая настройка волосковых клеток
Уши немлекопитающих позвоночных также частотно избирательны, несмотря на то, что у них базилярные мембраны не расширяются и не поддерживают механически настроенные бегущие волны. Кроуфорд и Феттиплейс предприняли исследования на черепахе, чтобы показать, что механосенсорные волосковые клетки обладают встроенной частотной избирательностью благодаря механизму электрической настройки80). Внутриклеточные отведения от волосковой клетки базилярного сосочка черепахи (слуховой сенсорный эпителий) показаны на рис. 18.16. Когда подавался короткий звуковой стимул (щелчок), возникали затухающие колебания мембранного