

Nikolls_-_Ot_neyrona_k_mozgu
.pdfГлава 11. Высвобождение медиатора |
213 |
Постсинаптический потенциал действия также постепенно уменьшается и в определенный момент полностью исчезает, так как возбуждающий постсинаптический потенциал (ВПСП) перестает достигать порога потенциала действия. С этого момента по амплитуде синаптического потенциала можно судить о количестве высвобождаемого медиатора.
График зависимости амплитуды возбуждающего постсинаптического потенциала от амплитуды пресинаптического импульса представлен на рис. 11.1C. Синаптический потенциал уменьшается при снижении амплитуды пресинаптического потенциала действия менее 75 мВ, а если амплитуда пресинаптического импульса становится меньше 45 мВ, то Постсинаптический ответ вовсе пропадает. Тетродотоксин не влияет на чувствительность постсинаптической мембраны к медиатору; поэтому снижение амплитуды синаптического потенциала связано с уменьшением количества медиатора, выделяемого из пресинаптического окончания. Таким образом, деполяризация на 45 мВ является пороговым значением для высвобождения медиатора; при еще большем увеличении амплитуды пресинаптического потенциала действия возрастают количество высвобождаемого медиатора и амплитуда ВПСП.
Для того, чтобы исследовать эту зависимость в деталях, Катц и Миледи провели следующий эксперимент: они ввели в пресинаптическое окончание второй электрод, через который подавались короткие (1-2 мс) деполяризующие толчки тока с целью имитировать пресинаптический потенциал действия. Зависимость амплитуды синаптического потенциала от амплитуды симулируемых пресинаптических потенциалов действия оказалась такой же, как и в случае настоящих потенциалов действия (см. рис. 11.1С). Этот результат свидетельствует о том, что ионные потоки натрия и калия, ответственные за генерацию потенциала действия, не являются обязательным фактором для высвобождения медиатора и что процесс высвобождения запускается деполяризацией.
Синаптическая задержка
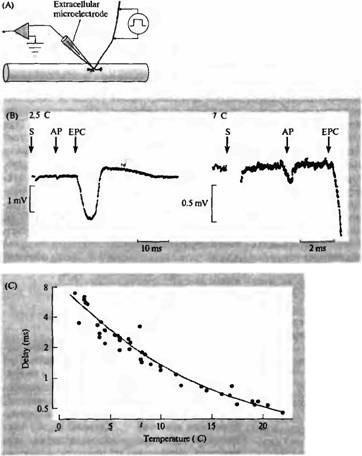
Одной из характерных особенностей процесса высвобождения медиатора является синаптическая задержка — время между началом пресинаптического потенциала действия и началом синаптического потенциала (рис. 11.1В; глава 9). В приведенных выше экспериментах на гигантском аксоне кальмара, которые проводились при температуре около 10° С, задержка составляла около 3-4 мс. В нервно-мышечном соединении лягушки синаптичес-кая задержка составляет 0,5 мс при комнатной температуре (рис. 11.2)7'. Эта задержка значительно превышает время (около 50 мкс), необходимое для диффузии АХ через синапти-ческую шель (расстояние около 50 нм). При ионофоретической аппликации АХ на постси-наптическую мембрану из пипетки задержка может составлять всего 150 мс. Более того, синаптическая задержка гораздо более чувствительна к температуре, чем процесс диффузии. Охлаждение нервно-мышечного препарата до 2° С увеличивает задержку до 7 мс (рис. 11.2В), в то время как задержка на ответ, вызываемый ионофоретической аппликацией АХ, значительно не меняется. Таким образом, основная причина задержки кроется в механизме высвобождения медиатора8).
Значение кальция для процесса высвобождения
Кальций был давно известен как важное звено в процессе синаптической передачи. При уменьшении концентрации кальция в наружном растворе уменьшается также и высвобождение АХ в нервно-мышечном соединении, вплоть до полного блокирования постсинаптических ответов9·|0). Кальций играет важную роль в высвобождении медиатора во всех синапсах независимо от природы медиатора (одним из исключений является высвобождение ГАМК из горизонтальных клеток в сетчатке рыб, см. главу 4")). Было также показано, что кальций играет важную роль в процессах секреции в целом, как, например, в высвобождении гормонов клетками гипофиза, выделении норадреналина из мозгового вещества надпочечников и секреции из слюноотделительных желез 12, 13). Как будет показано в следующем разделе, вызванному высвобождению медиатора предшествует вход кальция в пресинаптическое окончание, причем высвобождение может быть заблокировано ионами, которые препятствуют вхождению кальция, например, магнием, кадмием, никелем, марганцем и кобальтом. Следовательно, высвобождение медиатора можно уменьшить, либо устранив кальций из наружного раствора, либо добавив в него блокирующие ионы. Медиатор высвобождается только в том случае, если
214 |
|
|
|
|
|
|
|
Раздел II. Передача информации в нервной системе |
Рис. |
11.2. |
Синаптическая |
|
|||||
задержка |
|
в |
химическом |
|
||||
синапсе. |
|
(А) |
|
Ответы, |
|
|||
вызываемые |
|
|
|
|
|
|||
электрической |
|
|
|
|
||||
стимуляцией |
|
|
|
|
|
|||
двигательного |
|
нерва, |
|
|||||
регистрируются |
|
|
с |
|
||||
помощью |
|
внеклеточного |
|
|||||
микроэлектрода в нервно-- |
|
|||||||
мышечном |
|
соединении |
|
|||||
лягушки. |
|
|
|
|
|
(В) |
|
|
Экстраклеточная |
|
|
|
|
||||
регистрация |
|
артефакта |
|
|||||
стимуляции (S), |
потенциал |
|
||||||
действия окончания аксона |
|
|||||||
(АР) и тока концевой |
|
|||||||
пластинки (ЕРС) при 2,5 и |
|
|||||||
7° |
С. |
|
Синаптическая |
|
||||
задержка |
|
— это |
время |
|
||||
между |
|
|
потенциалом |
|
||||
действия |
|
|
в |
нервном |
|
|||
окончании и началом тока |
|
|||||||
концевой |
|
|
|
пластинки. |
|
|||
Обратите |
|
внимание, |
что |
|
||||
|
|
|||||||
ток, входящий в нервное |
|
|||||||
окончание |
и |
мышечное |
|
|||||
волокно, |
|
регистрируется |
|
|||||
внеклеточным |
|
|
|
|
||||
микроэлектродом |
в |
виде |
|
|||||
отрицательных |
|
|
|
|
||||
потенциалов. (С) График |
|
|||||||
зависимости |
|
|
|
|
|
|||
синаптической задержки от |
|
|||||||
температуры; |
чем |
выше |
|
|||||
температура, |
тем |
короче |
|
|||||
синаптическая задержка. |
|
|||||||
Fig. 11.2. Synaptic Delay at a |
|
|||||||
Chemical |
Synapse. (A) |
The |
|
|||||
motor |
nerve |
is |
stimulated |
|
||||
while |
recording |
with |
an |
|
||||
extracellular |
|
microelectrode |
|
|||||
|
|
|||||||
at the frog |
neuromuscular |
2,5 and 7° C. The synaptic delay is the time between the action potential in the nerve terminal |
||||||
junction. |
(B) |
Extracellular |
|
|||||
and the beginning of the end-plate current. Note that the current flowing into the nerve terminal |
||||||||
recordings |
|
of |
the |
stimulus |
or the muscle fiber is recorded as a negative potential by an extracellular microelectrode. (C) |
|||
artifact (S), the axon terminal |
Plot of synaptic delay as a function of temperature; the higher the temperature, the briefer the |
|||||||
action potential (AP), and the |
synaptic delay. (After Katz and Miledi, 1965.) |
|||||||
end-plate current (EPC) at |
|
|||||||
кальций присутствует в наружном растворе в момент деполяризации пресинаптического окончания 14).
Измерение входа кальция в пресинаптическое нервное окончание
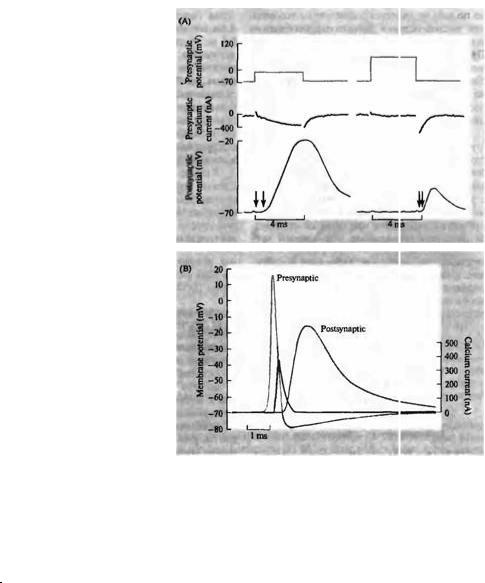
В ходе дальнейших экспериментов было выявлено, что проводимость мембраны для ионов кальция увеличивается при деполяризации и что кальций входит в окончание с каждым потенциалом действия. Ллинас с сотрудниками измерили амплитуду и временной ход кальциевого тока, вызываемого пресинаптической деполяризацией в гигантском синапсе кальмара. Пример такого эксперимента показан на рис. 11.3А. Натриевая и калиевая проводимости, связанные с потенциалом действия, были заблокированы ТТХ и ТЭА (тетраэтиламмонием), таким образом, только потенциалзависимые кальциевые каналы оставались функциональными. Деполяризация пресинаптического окончания до —18 мВ
Глава 11. Высвобождение медиатора |
215 |
|
||||||||||
Рис. 11.3. |
Пресинаптический кальций и |
|
|
|
||||||||
высвобождение медиатора в гигантском |
|
|
|
|||||||||
синапсе кальмара (А, В). Потенциал на |
|
|
|
|||||||||
лресиналтическом |
|
|
|
|
окончании |
|
|
|
||||
фиксируется на заданном уровне, а |
|
|
|
|||||||||
потенциал |
активируемые |
натриевые и |
|
|
|
|||||||
калиевые |
токи |
|
заблокированы |
те |
|
|
|
|||||
тродотоксином и тетраэтиламмонием. (А) |
|
|
|
|||||||||
На верхних записях показан командный |
|
|
|
|||||||||
потенциал, |
прикладываемый |
к |
|
|
|
|||||||
пресиналтическим волокнам, в середине |
|
|
|
|||||||||
— пресиналтический кальциевый |
ток, |
|
|
|
||||||||
внизу |
— |
|
|
возбуждающий |
|
|
|
|||||
постсиналтический потенциал (ВПСП) в |
|
|
|
|||||||||
постсинаптическом |
волокне. |
Толчок |
|
|
|
|||||||
потенциала от -70 до -18 мВ (слева) |
|
|
|
|||||||||
вызывает |
медленный |
|
|
входящий |
|
|
|
|||||
кальциевый ток, за которым с задержкой |
|
|
|
|||||||||
около 1 мс (отмеченной стрелками) |
|
|
|
|||||||||
следует ВПСП. Деполяризация до +60 мВ |
|
|
|
|||||||||
(справа) подавляет вход кальция. В конце |
|
|
|
|||||||||
импульса возникает |
волна |
кальциевого |
|
|
|
|||||||
тока, за которой с задержкой около 0,2 мс |
|
|
|
|||||||||
следует ВПСП. (В) Если в условиях |
|
|
|
|||||||||
фиксации потенциала вызвать такое же |
|
|
|
|||||||||
по |
форме |
изменение |
|
мембранного |
|
|
|
|||||
потенциала, как во время нормального |
|
|
|
|||||||||
потенциала |
действия |
|
|
(отметка |
|
|
|
|||||
Presynaptic), то возникающий при этом |
|
|
|
|||||||||
ВПСП невозможно отличить от ВПСП, |
|
|
|
|||||||||
возникающего |
в |
физиологических |
|
|
|
|||||||
условиях |
(отметка |
|
Post-synaptic). |
|
|
|
||||||
Амплитуда и временной ход кальциевого |
|
|
|
|||||||||
тока |
показаны |
|
черной |
|
линией. |
|
|
|
||||
Синалтическая задержка между началом |
|
|
|
|||||||||
пресиналтической |
|
деполяризации |
и |
|
|
|
||||||
началом |
лостсинаптического |
ответа |
|
|
|
|||||||
отчасти |
обусловлена |
|
временем, |
|
|
|
||||||
затрачиваемым |
|
на |
|
|
открывание |
|
|
|
||||
кальциевых каналов, а также на то, чтобы |
|
|
|
|||||||||
вошедший |
кальций |
|
|
запустил |
|
|
|
|||||
высвобождение медиатора. |
|
|
|
|
|
|
|
|||||
|
|
|
|
|
-70 to -18 mV (left panel) results in a slow inward calcium current and, after a delay |
|
||||||
Fig. |
11.3. |
Presynaptic |
|
Calcium |
and |
|
|
|||||
|
of about 1 ms (arrows), an EPSP. A larger depolarization, to +60 mV (right panel), |
|||||||||||
Transmitter |
Release |
at |
the |
squid |
giant |
|
||||||
synapse. (A, B) The presynaptic terminal is |
|
suppresses calcium entry. At the end of the pulse, a surge of calcium current is |
||||||||||
voltage-clamped and treated with TTX and |
|
followed within about 0,2 ms (arrows) by an EPSP. (B) If a voltage change identical |
||||||||||
TEA to abolish voltage-activated sodium |
|
in shape to a normal action potential is produced by the voltage clamp (labeled |
||||||||||
and potassium currents (A). Records show |
|
Presynaptic), then the EPSP is indistinguishable from that seen normally (labeled |
||||||||||
potentials appb'ed |
to |
the |
presynaptic |
fiber |
|
Postsynaptic). The black curve gives the magnitude and time course of the calcium |
||||||
(upper trace), presynaptic calcium current |
|
current. The synaptic delay between the beginning of presynaptic depolarization |
||||||||||
(middle trace), and EPSP in the postsynaptic |
|
and the beginning of postsynaptic response is due in part to the time required to |
||||||||||
fiber (lower trace). A voltage pulse from |
|
|
open calcium channels and in part to the time for calcium entry to trigger |
|||||||||
|
|
transmitter release. (After Llinàs, 1982.) |
||||||||||
|
|
|
|
|
|
|
|
|
|
|
||
(верхняя запись слева) вызывала входящий кальциевый ток в окончание, который медленно увеличивался примерно до 400 нА (средняя запись), а также синаптический потенциал большой амплитуды в постсинаптической клетке (нижняя запись). Если окончание деполяризовалось до +60 мВ, что примерно соответствует равновесному потенциалу кальция, то кальциевый ток и синаптический потенциал подавлялись во время деполя-

216 |
Раздел II. Передача информации в нервной системе |
ризационного толчка (см. записи справа). Это доказывает, что деполяризация окончания сама по себе не является достаточным условием для высвобождения медиатора; необходимо также, чтобы произошел вход ионов кальция. При реполяризации наблюдался кратковременный кальциевый ток, как если бы кальций входил через каналы, которые открылись во время деполяризации, и это сопровождалось небольшим постсинаптическим потенциалом.
Эффект искусственного потенциала действия показан на рис. 11.3В. Пресинаптический потенциал действия, записанный до добавления ТТХ и ТЭА, был «воспроизведен» электрической цепью фиксации потенциала, с тем чтобы вызвать точно такое же изменение потенциала в окончании. Постсинаптический потенциал при этом не отличается от ответа, вызванного нормальным пресинаптическим потенциалом действия, подтверждая тот факт, что натриевые и калиевые токи, которые в норме участвуют в генерации потенциала действия, не являются необходимым условием для высвобождения медиатора.
Использование методики фиксации потенциала также позволило измерить амплитуду и временной ход кальциевого тока, вызываемого искусственным потенциалом действия (кривая черного цвета на рис. 11.3В). Кальциевый ток начинается примерно через 0,5 мс после начала пресинаптической деполяризации, а постсинаптический потенциал начинается еще через 0,5 мс. Таким образом, время, необходимое для деполяризации пресинаптического окончания и для открывания кальциевых каналов, составляет первую половину синаптической задержки; оставшаяся часть задержки уходит на возрастание концентрации кальция в окончании и высвобождение медиатора.
Ллинас с сотрудниками также непосредственно наблюдали вход кальция, используя люминесцентную метку экворин 15· 16). Они обнаружили, что в результате короткой серии пресинаптических потенциалов действия внутриклеточная концентрация кальция достигает от 100 до 200 мкмоль в микродоменах внутри окончания (рис. 11.4), которые, по всей видимости, соответствуют активным зонам (глава 9).
Локализация мест входа кальция
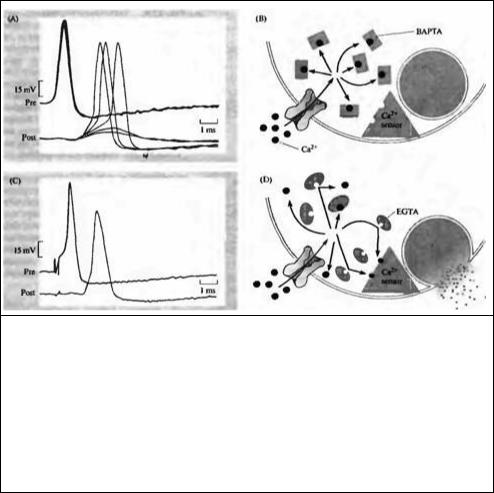
Эксперименты на гигантском синапсе кальмара дали дополнительную информацию о роли кальция в высвобождении и, в частности, о близком расположении кальциевых каналов к местам секреции медиатора 17). В этих экспериментах введение ВАРГА — сильного кальциевого буфера — в пресинаптическое окончание вызывало значительное уменьшение количества высвобождаемого медиатора без какого-либо существенного изменения потенциала действия (рис. 11.5А). С другой стороны EGTA — другой кальциевый буфер — обладал лишь небольшим эффектом на высвобождение (рис. 11.5С). Это различие связано с тем, что связывание кальция с ВАРТА происходит намного быстрее, чем с EGTA. Таким образом, у кальциевых ионов остается незначительная возможность для диффузии от места входа, прежде чем они свяжутся с ВАРТА; в то же время ионы кальция могут проходить более значительные расстояния, прежде чем будут связаны с EGTA (рис. 11.5В и D). По скорости диффузии кальция и его связыванию с EGTA можно рассчитать, что место связывания кальция, связанное с процессом высвобождения, должно находиться в пределах 100 нм от места входа кальция. С другой стороны, было показано, что EGTA
Рис. 11.4. (Рис. 11.4А и 11.4В см. на цветной вклейке после с. 640.) Микродомены кальция в пресина птическом окончании в гигантском синапсе кальмара. (А) Распределение кальция внутри пресинаптическо го окончания аксона в покое, определенное с помощью внутриклеточной инъекции кальций-чувствительной метки (квадратом на рисунке слева отмечен участок, в котором проводится измерение кальция). (В) Короткая серия пресиналтических потенциалов действия вызывает появление микродоменов с высокой концентрацией кальция внутри окончания аксона.
Fig. 11.4. Microdomains of Calcium within the Presynap tic Terminal at the Squid Giant Synapse. (A) Distribution of calcium within the presynaptic axon terminal at rest determined by intracellular injection of a calcium-sensitive dye (box in illustration at leftj shows the region imaged). (B) A brief train of presynaptic action potentials results in the appearance of microdomains of high calcium concentration within the axon terminal (After Llinas, Sugimori, and Silver, 1992; micrographs kindly provided by R. Llinas.)
Глава И. Высвобождение медиатора |
217 |
|
|
Рис. 11.5. Кальций входит около мест высвобождения медиатора в гигантском синапсе кальмара. (А) Внутриклеточная регистрация от лре- и постсинаптического аксонов после инъекции быстрого кальциевого хелатора ВАРГА. Наложенные друг на друга записи показывают уменьшение ВПСП в течение 4-х минут после инъекции ВАРГА. (В) ВАРГА связывает кальций до того, как он достигает «кальциевого сенсора», который запускает высвобождение медиатора. (С) Наложенные друг на друга внутриклеточные записи, полученные в течение 4-х минут инъекции ЕСГА — хелатора, который связывает кальций гораздо медленнее. Амплитуда ВПСП не изменяется. (D) Кальций достигает «кальциевого сенсора», который запускает высвобождение медиатора, быстрее, чем с ним свяжется ЕСГА; это указывает на то, что место входа кальция должно быть в пределах 100 нм от места, в котором кальций запускает высвобождение медиатора.
Fig. 11.5. Calcium Enters Near the Site of Transmitter Release at the squid giant synapse. (A) Intracellular recordings from the preand postsynaptic axons following injection of the fast calcium chelator ВАРГА. Superimposed traces show the reduction in the EPSP during a 4 min ВАРТА injection. (В) ВАРГА binds calcium before it has time to reach the calcium sensor that triggers release. (C) Superimposed intracellular recordings during a 4 min injection of ЕСГА, a chelator that binds calcium more slowly. No change in EPSP amplitude is seen. (D) Calcium reaches the sensor that triggers release faster than it becomes bound to ЕСГА, indicating that the site of calcium entry must be within 100 nm of the site at which calcium triggers transmitter release. (A and С after Adler et al., 1991.)
может влиять на высвобождение медиатора в некоторых синапсах; это предполагает, что в этих клетках кальций может диффундировать на большие расстояния от кальциевых каналов до мест, в которых инициирется высвобождение медиатора5).
Роль деполяризации в высвобождении медиатора
Представленные до сих пор доказательства указывают на то, что высвобождение медиатора запускается увеличением внутриклеточной концентрации ионов кальция, которая происходит при деполяризации пресинаптического окончания и открывании потенциалзависимых кальциевых каналов. Данная гипотеза была проверена с помощью так называемого "caged calcium" — буфера, который высвобождает кальций при облучении ультрафиолетовым светом 18) -- 20). При инъекции нитрофена (одна из форм "caged calcium") в пресинаптическое окончание гигантского синапса кальмара или нервно-мышечное соединение рака облучение окончания вызывало высвобождение медиатора (рис. 11.6). Для высвобождения такого же количества медиатора, как и при синаптическом потенциале

218 |
Раздел II. Передача информации в нервной системе |
Рис. 11.6. Увеличение внутриклеточного кальция достаточно для того, чтобы вызвать быстрое высвобождение медиатора в гигантском синапсе кальмара. (А) Нитрофен (вещество, которое высвобождает кальций при облучении ультрафиолетом) инъецирован в пресиналтическое окончание. Высвобождение медиатора регистрируется с помощью внутриклеточного отведения от постсинаптического аксона. (В) Внутриклеточные постсинаптические ответы, вызванные стимуляцией нерва (ВПСП) и высвобождением кальция из нитрофена под действием вспышки ультрафиолетового света
(nitrophen response). Резкое увеличение внутриклеточного кальция вызывает увеличение освобождения медиатора почти такое же быстрое, как при прохождении пресинаптического потенциала действия. Спад ответа, вызванного нитрофеном, медленный и неполный, поскольку после фотолиза нитрофен удерживает концентрацию кальция на более высоком уровне, чем в нормальных условиях.
Fig. 11.6. An Increase in IntraceUuLar Calcium Is Sufficient to Trigger Rapid Transmitter Release at the squid giant synapse. (A) Nitrophen, a form of "caged calcium," is injected into the presynaptic terminal. Transmitter release is monitored by recording intracellularly from the postsynaptic axon. (B) Intracellular records show the postsynaptic response to nerve stimulation (EPSP) and to release of calcium from nitrophen by a flash of ultraviolet light (nitrophen response). An abrupt increase in intracellular calcium causes an increase in transmitter release that is nearly as rapid as that produced by a presynaptic action potential. The decay of the nitrophen response is
slower and incomplete because the photolyzed nitrophen buffers calcium to a concentration higher than the normal level at rest. (B after Zucker, 1993.) ·
действия, было необходимо, чтобы внутриклеточная концентрация кальция возрастала примерно до 100 мкмоль. В экспериментах на окончаниях биполярных клеток из сетчатки золотой рыбки были получены схожие результаты 21).
В тоже время некоторые свойства процесса высвобождения не могут быть объяснены лишь на основании входа кальция в нервное окончание. Эти свойства были исследованы в нервномышечных синапсах рака и лягушки, и было обнаружено, что при удержании внутриклеточной концентрации кальция на постоянном повышенном уровне и блокировании дополнительного входа кальция деполяризация может вызывать высвобождение медиатора 8). Место действия деполяризации не установлено; одним из объяснений является то, что деполяризация влияет на потенциалзависимые ауторецепторы в мембране пресинаптического нервного окончания 22).
§ 2. Квантовое высвобождение медиатора
Исходя из вышесказанного, общая схема высвобождения медиатора может быть представлена в виде следующей цепочки:
Теперь остается показать, каким образом медиатор высвобождается из нервных окончаний. В экспериментах на нервно-мышечном соединении лягушки Фетт и Катц показали, что АХ может высвобождаться из окончаний в виде мультимолекулярных пакетов «квантов» 23). Последующие эксперименты показали, что каждый квант соответствует при-
Глава 11. Высвобождение медиатора |
219 |
|
|
||||||||
Рис. |
11.7. |
Миниатюрные |
|
|
|
|
|
||||
синалтические |
|
|
|
|
|
|
|
|
|||
потенциалы |
|
возникают |
|
|
|
|
|
||||
спонтанно |
|
в |
нервно- |
|
|
|
|
|
|||
мышечном |
|
соединении |
|
|
|
|
|
||||
лягушки. |
|
|
|
(А) |
|
|
|
|
|
||
Внутриклеточная |
|
|
|
|
|
|
|
||||
регистрация |
|
мышечного |
|
|
|
|
|
||||
волокна |
|
в |
|
области |
|
|
|
|
|
||
двигательной |
|
|
концевой |
|
|
|
|
|
|||
пластинки. (В) Спонтанные |
|
|
|
|
|
||||||
миниатюрные |
|
|
|
|
|
|
|
|
|||
синалтические |
потенциалы |
|
|
|
|
|
|||||
имеют |
амплитуду |
около 1 |
|
|
|
|
|
||||
мВ |
|
и |
|
наблюдаются |
|
|
|
|
|
||
исключительно |
в |
области |
|
|
|
|
|
||||
концевой пластинки. (С) |
|
|
|
|
|
||||||
После |
|
|
|
добавления |
|
|
|
|
|
||
лростигмина |
|
(ингибитора |
|
|
|
|
|
||||
фермента |
|
|
|
|
|
|
|
|
|
||
ацетилхолинэстеразы, |
|
|
|
|
|
|
|||||
которая |
гидролизует |
АХ) |
|
|
|
|
|
||||
миниатюрные |
|
|
|
|
|
|
|
|
|||
синаптические |
потенциалы |
|
|
|
|
|
|||||
увеличиваются |
в амплитуде |
|
|
|
|
|
|||||
и длительности, но при |
|
|
|
|
|
||||||
этом |
|
их |
|
частота |
не |
|
|
|
|
|
|
изменяется. |
Это |
указывает |
|
|
|
|
|
||||
на |
то, |
что |
каждый |
|
|
|
|
|
|||
миниатюрный |
потенциал |
|
|
|
|
|
|||||
является |
|
ответом |
на |
|
|
|
|
|
|||
квантовую порцию АХ, а не |
|
|
|
|
|
||||||
|
|
|
|
||||||||
ответом |
на |
одну |
молекулу |
|
recording from a muscle fiber in the region of the motor end plate. (B) |
Spontaneous miniature |
|
||||
АХ. |
|
|
|
|
|
|
|
synaptic potentials are about 1 mV in amplitude and are confined to the |
end-plate region of the |
||
Fig. 11.7. Miniature Synaptic |
|
muscle fiber. (C) After addition of prostigmine, which prevents |
acetylcholinesterase from |
||||||||
|
hydrolyzing ACh, miniature synaptic potentials are increased in amplitude |
and duration, but the |
|||||||||
Potentials occur spontaneously |
|
frequency at which they occur is unchanged. This indicates that each miniature is due to a quantal |
|||||||||
at the |
frog |
|
neuromuscular |
|
packet of ACh, rather than to a single ACh molecule. (After Fatt and Katz, 1952.) |
||||||
junction. (A) Intracellular |
|
|
|
|
|
|
|||||
мерно 7000 молекул АХ 24). Следовательно, квантовое высвобождение означает, что только 0, 7000, 14 000 и так далее молекул может высвобождаться одновременно, а не 4 250 или 10 776. В каждом данном синапсе количество квантов, высвобождаемых из нервного окончания в ответ на потенциал действия (квантовый состав синаптического ответа) может значительно варьировать, однако количество молекул в каждом кванте (размер кванта) является постоянной величиной (с вариациями в пределах 10%).
Спонтанное высвобождение квантов медиатора
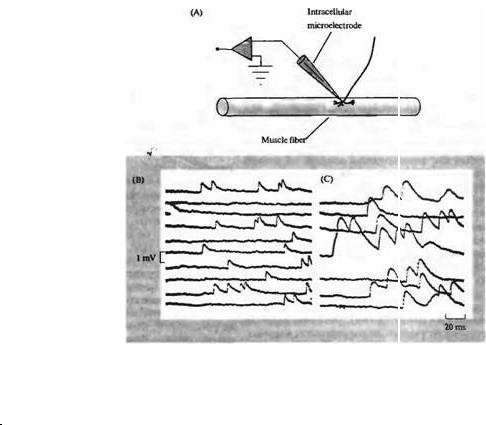
Первым свидетельством того, что АХ упакован в мультимолекулярные кванты, было то, что в области концевой пластинки наблюдается спонтанная нерегулярная деполяризация с амплитудой около 1 мВ (рис. 11.7) 23). Временной ход спонтанных деполяризаций совпадал с временным ходом потенциала, вызываемого стимуляцией нерва. Спонтанные миниатюрные потенциалы (МПКП, миниатюрные потенциалы концевой пластинки) уменьшались по амплитуде вплоть до полного исчезновения по мере увеличения концентрации антагониста АХ рецептора кураре, в то время как ингибиторы ацетилинхолинэстеразы (например, простигмин) увеличивали амплитуду и замедляли временной ход МПКП (рис. 11.7С). Эти два фармакологических теста указывают на то, что МПКП генерируется в результате спонтанного высвобождения дискретных количеств АХ из нервного окончания, и исключают возможность того, что МПКП генерируется одиночными молекулами АХ. Впоследствии регистрация с помощью пэтч-электродов позволила непосредственно определить, что количество тока, протекающего через один АХ рецептор, должно вызывать деполяризацию мышечного
220 |
Раздел II. Передача информации в нервной системе |
волокна приблизительно на 1 мкВ (глава 2). Следовательно, в генерации каждого спонтанного МПКП участвует более тысячи АХ рецепторов. Были получены также дополнительные доказательства того, что спонтанные миниатюрные потенциалы являются ответом на мультимолекулярные пакеты АХ, выделяемые нервным окончанием. Например, деполяризация нервного окончания, вызываемая постоянным током, приводит к увеличению частоты спонтанной активности, в то время как деполяризация мышцы не влияет на частоту25'. Ботулиновый токсин, который блокирует высвобождение АХ в ответ на стимуляцию нерва, блокирует также и спонтанную активность26'. Вскоре после денервации мышцы двигательное нервное окончание дегенерирует, что сопровождается исчезновением миниатюрных потенциалов27'. Интересно, что по прошествии короткого промежутка времени спонтанные потенциалы вновь появляются в денервированной мышце лягушки; они возникают в результате высвобождения АХ шБанковскими клетками, которые поглощают участки дегенерирующих нервных окончаний путем фагоцитоза28'.
Неквантовое высвобождение
Вдополнение к квантовой форме высвобождения, некоторое количество АХ также
постоянно «утекает» из цитоплазмы во внеклеточное пространство в неквантовой форме29·30). В действительности, общее количество АХ, который утекает из нервного окончания в виде неквантового высвобождения, примерно в 100 раз больше, чем количество АХ, выделяемого в виде спонтанных квантов. Это соотношение может быть определено путем сравнения общего количества высвобождаемого из мышцы АХ, измеренного биохимическими методами, с количеством АХ, высвобождаемого в виде квантов, рассчитанным по частоте МПКП с учетом общего количества концевых пластинок на мышце.
Вфизиологических условиях утечка АХ из пресинаптических окончаний в неквантовой форме не вызывает постсинаптического ответа: количество холинэстеразы в синаптической щели достаточно для того, чтобы гидролизовать столь незначительное количество АХ, предотвращая таким образом активацию рецепторов на постсинаптической мембране. Постсинаптический эффект неквантового высвобождения может быть выявлен только в условиях ингибированной холинэстеразы. В то же время одновременное высвобождение 7 000 молекул АХ в одном кванте локально преодолевает активность этого фермента за счет субстратного ингибирования и позволяет АХ достичь постсинаптических рецепторов и вызвать МПКП (глава 13).
Флуктуации потенциала концевой пластинки
Типичный синаптический потенциал в скелетном нервно-мышечном соединении деполяризует постсинаптическую мембрану на 50--70 мВ. Какой механизм высвобождения АХ — утечка или кванты — лежит в основе этого синаптического потенциала? Фетт и Катц обнаружили, что при уменьшении синаптической передачи путем понижения экстраклеточной концентрации кальция и увеличения концентрации магния постсинаптические ответы, вызываемые стимуляцией нервов, флуктуируют ступенчатым образом, как показано на рис. 11.8А23'. Некоторые стимулы не вызывают ответов вовсе, некоторые вызывают ответы с амплитудой около 1 мВ, идентичные по размеру и форме спонтанным миниатюрным потенциалам; наконец, остальные вызванные ответы имеют двух-, трех--или четырехкратную амплитуду по отношению к амплитуде миниатюрных спонтанных потенциалов.
Это удивительное наблюдение привело к квантовой гипотезе, в которой предполагается, что одиночные квантовые события, возникающие спонтанным образом, представляют собой также «строительные блоки» для синаптических потенциалов, вызванных стимуляцией. В физиологических условиях потенциалы концевой пластинки состоят примерно из 200 квантовых единиц, и вариации в их амплитуде незначительны. В условиях пониженной концентрации кальция размер кванта остается неизменным, однако квантовый состав очень мал — около 1, 2 или 3 квантов — и случайным образом флуктуирует от одного события к другому, приводя к ступенеобразным флуктуациям в амплитуде потенциала концевой пластинки.
Статистический анализ потенциалов концевой пластинки
Кастильо и Катц решили применить статистический анализ для проверки квантовой

Глава 11. Высвобождение медиатора |
221 |
|
|
Рис. 11.8. Потенциалы концевой пластинки состоят из квантовых единиц, которые соответствуют спонтанным миниатюрным потенциалам. Пресинаптическое высвобождение АХ в нервно-мышечном окончании лягушки было уменьшено за счет снижения концентрации кальция в наружном растворе. (А) Примеры ответов, вызываемых стимуляцией нерва и регистрируемых внутриклеточно от мышечного волокна. Амплитуда потенциалов концевой пластинки (ПКП) варьирует ступенчатым образом; самые маленькие ответы по амплитуде соответствуют спонтанным миниатюрным потенциалам (МПКП). (В) Сравнение среднего квантового состава (m) ПКП, определенного двумя способами: применением распределения Пуассона, m — In (N/n0), и путем деления средней амплитуды ПКП на среднюю амплитуду МПКП. Великолепное соответствие этих двух оценок подтверждает гипотезу о том, что ПКП состоят из квантовых единиц, соответствующих спонтанным МПКП.
Fig. 11.8. The End-Plate Potential Is Composed of Quantal Units That Correspond to Spontaneous Miniature Potentials. Presynaptic release of ACh at a frog neuromuscular junction was reduced by lowering the calcium concentration in the bathing solution. (A) Sets of intracellular records, each showing two to four superimposed responses to nerve stimulation. The amplitude of the end-plate potential (EPP) varies in a stepwise fashion; the smallest response corresponds in amplitude to a spontaneous miniature potential (MEPP). (B) Comparison of the mean quantal content (m) of the EPP determined in two ways: by applying the Poisson distribution, m = In (N/n0). and by dividing the mean EPP amplitude by the mean MEPP amplitude. Agreement of the two estimates supports the hypothesis that the EPP is composed of quantal units that correspond to spontaneous MEPPs. (A after Fatt and Katz, 1952; В after del CastiUo and Katz, 1954.)
гипотезы 31). Они предположили, что двигательное нервное окончание содержит тысячи квантовых пакетов АХ (n), каждый из которых имеет вероятность (р) быть высвобожденным в ответ на нервный импульс, а также что кванты высвобождаются независимо, т. е. высвобождение одного не влияет на вероятность высвобождения последующего. При большом количестве событий среднее количество квантов (m), высвобождаемых во время события, будет равным пр, а количество событий, состоящих из 0, 1, 2, 3, 4,... или x квантов, должно соответствовать биномиальному распределению. Однако Кастильо и Катц не имели возможности измерить n или p экспериментально, и поэтому они не могли использовать биномиальное распределение для проверки гипотезы о том, что потенциал концевой пластинки состоит из единиц одинакового размера, соответствующих спонтанным миниатюрным потенциалам. Для того, чтобы справиться с этой проблемой, они рассуждали следующим образом:
В нормальных условиях можно предположить, что ρ сравнительно велика, т. е. большая часть синоптической популяции отвечает на импульс. Однако как только мы уменьшаем концентрацию кальция и увеличиваем концентрацию магния, вероятность ответа уменьшается, и мы не наблюдаем ответов на большую часть стимулов, и редко — ответы, состоящие из одной или двух единиц. В этих условиях, когда ρ очень нала, количество единиц х, которые составляют ПКП, при большом количестве наблюдений должно распределяться по закону Пуассона.
Распределение Пуассона является приближением к биномиальному распределению в случаях малых значений р. Принципиальным отличием является то, что для предсказания Пуассоновского распределения не обязатель-

222 |
Раздел II. Передача информации в нервной системе |
но знать n или р. Экспериментатору необходимо измерить лишь их общий продукт m, среднее количество квантов, высвобождаемых во время одного события. Предполагаемое количество ответов, состоящих из x квантов, задается в распределении Пуассона по формуле:
Одним из лучших примеров использования распределения Пуассона в истории был анализ числа прусских кавалерийских офицеров, убиваемых за год ударом лошадиного копыта. При большом количестве офицеров (n) вероятность (р) того, что кто-то из них будет убит, очень мала. В некоторые годы никто из офицеров не погибал; в остальные погибали один, а иногда и двое. В течение длительного периода наблюдений количество лет, в которые погибали 0, 1, 2 или 3 офицера, приближались к значениям из уравнения Пуассона, при использовании только средних значений «удачных» пиков в год (m) для определения теоретически предполагаемого распределения.
Другим примером, в котором уравнение Пуассона может предсказать распределение событий, является игровой автомат типа «однорукий бандит». Размер одной единицы равен 5 центам, автомат содержит большое количество монет, а вероятность выпадения каждой монеты очень мала и не зависит от других монет. Если известно среднее количество монет, выплачиваемых за игру, то при длительном наблюдении за игрой уравнение Пуассона точно предскажет, сколько раз игра будет проиграна, а также сколько раз игрок получит одну, две и более монет. И вновь важным свойством уравнения Пуассона является то, что свойство распределения зависит только от m.
Следовательно, для того чтобы проверить, подчиняются ли флуктуации потенциалов концевой пластинки при сниженной концентрации кальция закону Пуассона, необходимым является лишь значение m, среднее количество единиц, высвобождаемых за одно событие. Это значение получается путем деления средней амплитуды вызванных потенциалов на размер единицы — среднюю амплитуду спонтанных миниатюрных потенциалов:
В случае игрового автомата m равняется среднему количеству денег, выплачиваемому по результатам каждой игры (по всей видимости, не очень много, скажем, 1,5 цента за игру), деленному на размер единицы (5 центов), что дает m = 0,3 единицы за игру. Если амплитуды потенциалов концевой пластинки распределяются в соответствии с уравнением Пуассона, то m также может быть определено по количеству нулевых ответов П0. В уравнении Пуассона при x = 0 n0 = Ne–m (поскольку m0 и 0! = 1). Таким образом, формула может быть упрощена до:
Кастильо и Катц провели регистрацию большого количества потенциалов концевой пластинки, вызванных стимуляцией нерва, в условиях пониженного кальция и повышенного магния в наружном растворе, а также большого количества спонтанных миниатюрных потенциалов. Рассчитав m этими двумя способами, они обнаружили практически полное совпадение результатов, что послужило серьезным доказательством в пользу квантовой гипотезы (рис. 11.8В).
С целью дальнейшей проверки квантовой гипотезы можно попытаться предсказать полное распределение амплитуд ответов, используя значение m и средней амплитуды миниатюрного потенциала (рис. 11.9). Как и ранее, m рассчитывается как соотношение между средним вызванным потенциалом и средним спонтанным миниатюрным потенциалом. Затем рассчитывается количество предполагаемых ответов, состоящих из 0, 1, 2, 3,... квантов. Для принятия в расчет небольшой вариации в амплитуде кванта предполагаемое количество ответов, содержащих один квант, распределяется от среднего размера кванта с тем же разбросом, как и у спонтанных событий (рис. 11.9, вставка). Предполагаемое количество ответов, состоящих из 2, 3 и более квантов, распределяется вокруг средних значений с пропорционально увеличивающимся разбросом. Отдельные распределения затем суммируются для получения теоретического распределения, показанного на рисунке сплошной линией. Согласие с экспериментально полученным распределением (столбики) обеспечивает дополнительное доказательство квантовой гипотезы.