

Nikolls_-_Ot_neyrona_k_mozgu
.pdfГлава б. Ионные механизмы потенциала действия |
103 |
Рис. 6.1. Роль натрия в генерации потенциала действия. Потенциал действия, отведенный от аксона кальмара в морской воде (черный сплошной график), в растворах, содержащих 50% (штрих-пунктир) и 33% (серый) натрия от нормы, и затем снова в морской воде (пунктир).
Fig. 6.1. Role of Sodium in Action Potential Generation. Action potentials recorded from a squid axon bathed in seawater (black solid line), in solutions containing 50% (dashdot line) and 33 % normal sodium (grey), and then returned to seawater (dashed line).
(After Hodgkin and Katz, 1949.)
медленнее, чем в действительности. Причина этого в том, что общая проводимость мембраны
впокое достаточно мала, и выход накопленных в результате потенциала действия положительных ионов из клетки занял бы несколько миллисекунд, а то и десятки миллисекунд. В действительности, возврат к уровню потенциала покоя происходит гораздо быстрее благодаря значительному увеличению другой мембранной проводимости, а именно, за счет открытия калиевых каналов. С той же быстротой, с которой мембранный потенциал деполяризовался в направлении ENa, он гиперполяризуется на фазе спада потенциала действия
внаправлении ЕK. Увеличение калиевой проводимости может продолжаться до нескольких миллисекунд, что приводит во многих клетках не только к реполяризации, но и к гиперполяризации мембраны до значений более отрицательных, чем потенциал покоя (рис. 6.1).
Обобщим вышесказанное. Потенциал действия является результатом быстрого и значительного увеличения натриевой проводимости мембраны. Вход большого количества ионов хлора и аккумуляция положительного заряда на внутренней поверхности мембраны клетки сдвигает мембранный потенциал в направлении ENa. Реполяризация мембраны происходит в результате последующего увеличения калиевой проводимости и уменьшения внутриклеточного положительного заряда из-за выхода ионов калия из клетки. Мембранный потенциал при этом вновь приближается к Ек. Такое объяснение механизма генерации потенциала действия ведет к пониманию принципов распространения импульса, рассматриваемых в главе 7.
Какое количество ионов входит в клетку и выходит из нее во время потенциала действия?
Если учесть, что на фазе роста потенциала действия в клетку входит большое количество натрия и большое количество калия ее покидает на фазе спада, то становится очевидным, что концентрации этих ионов в цитоплазме должны измениться. Величину этих изменений можно определить либо экспериментально, либо путем расчетов.
Вычисления соотношения между мембранным потенциалом и трансмембранных концентрационных градиентов приведены в главе 7. При потенциале -67 мВ на внутренней поверхности мембраны находится приблизительно 4 · 1011 отрицательных зарядов на см2. На пике потенциала действия (+40 мВ) вместо этого отрицательного заряда внутри клетки накапливается около 2,4 · 1011 положительных зарядов, что происходит в результате входа в клетку 6,4 · 1011 ионов натрия на см2. Это соответствует приблизительно 10–12 моль/см2. Экспериментальные измерения входа радиоактивного натрия и выхода радиоактивного калия4) дали значения между 3 · 10–12 и 4 · 10–12 моль/см2. Различие между теоретически предсказанным и экспериментальным значениями объясняются главным образом тем, что при расчетах не принималось во внимание то, что вход натрия и выход калия частично перекрываются во времени. Таким образом, количество входящего натрия превышает значение, необходимое для деполяризации мембраны до пикового уровня потенциала действия, поскольку выход калия начинается до момента достижения пика.
Каково влияние входа натрия на его внутриклеточную концентрацию? Сегмент ак-
104 |
Раздел II. Передача информации в нервной системе |
|
|
|
|
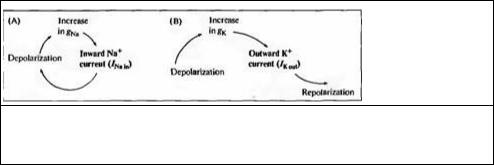
Рис. 6.2. Влияние повышения натриевой и калиевой проводимостей на мембранный потенциал. (А) Вход натрия усиливает деполяризацию. (В) Выход калия вызывает реполяризацию.
Fig. 6.2. Effects of Increasing Conductances (gNa and gK) on membrane potential. (A) Sodium entry reinforces depolarization. (B) Potassium efflux leads to repolarization.
сона кальмара длиной 1 см и диаметром 1 мм имеет площадь поверхности около 0,31 см2, поэтому при скорости входа натрия 3,5 · 10–12 приведет к накоплению приблизительно 10– 12 M натрия. Объем этого отрезка аксона составляет 7,85 · 10–12 л, в нем содержится (при плотности 50 ммоль/л) 4 · 10–7 M натрия, поэтому изменение концентрации натрия в результате потенциала действия очень мало и составляет 0,0000025. Выход калия производит сравнимый эффект и изменяет внутриклеточную концентрацию калия в
0,000003 раз.
Потенциал действия в более тонких отростках нейрона может привести к более значительным изменениям внутриклеточных концентраций натрия и калия, чем в гигантском аксоне кальмара. Так, нервное окончание диаметром 1 мкм и длиной 100 мкм имеет площадь поверхности 3 · 10–6 см2 и объем 8· 10–14 л. Во время потенциала действия вход натрия 3 · 10–6 моль/см2 производит увеличение количества натрия внутри терминали на 10–17 М. При внутриклеточной концентрации 20 мМ в терминали содержится приблизительно 1,5·10–15 М, так что увеличение концентрации натрия в результате одного потенциала действия составляет 0,7 %. Пачка из 50 импульсов теоретически приведет к увеличению уровня натрия на 35 % и сходному снижению уровня калия. Вход натрия увеличивает активность натрий-калиевого обменника (глава 4) так, что концентрации быстро возвращаются к изначальному уровню.
Положительная и отрицательная обратная связь во время изменений проводимости
Главным свойством натриевой и калиевой проводимостей, определяющим характер токов во время потенциала действия, является их потенциалзависимость: вероятность открытия ионных каналов увеличивается с деполяризацией мембранного потенциала. Деполяризация увеличивает натриевую проводимость, а также, с некоторой задержкой, калиевую. Воздействие деполяризации на натриевую проводимость носит регенеративный характер: сначала небольшая деполяризация увеличивает количество открытых каналов; ионы натрия, входящие в клетку по направлению своего электрохимического градиента, производят дальнейшую деполяризацию мембраны, открытие большего количества каналов, что влечет за собой вход еще большего числа ионов, и так далее (рис. 6.2А). Такой самоусиливающийся процесс характеризуется положительной обратной связью. Потенциал--зависимость калиевых каналов, наоборот, характеризуется наличием отрицательной обратной связи (рис. 6.2В). При деполяризации количество открытых калиевых каналов возрастает, и ионы калия входят в клетку в направлении электрохимического градиента. Однако, вход калия не усиливает деполяризацию, а ведет к реполяризации и возвращению калиевой проводимости в состояние покоя.
Измерения проводимости
То, что до сих пор обсуждалось в этой главе, было предложено Ходжкином, Хаксли и Катцем 5) и детально разработано Ходжкином и Хаксли6) – 9), которые поставили элегантные опыты на гигантском аксоне кальмара. Они экспериментально показали, что при потенциале действия происходят изменения калиевой и натриевой проводимостей, и что последовательность этих изменений в точности определяет временной ход и величину потенциала действия.
На основании каких опытов был сделан этот вывод? На первый взгляд, измерение натриевой (gNa) и калиевой (gк) проводимостей кажется простой задачей. Нужно только измерить величину тока (I), протекающего через

Глава 6. Ионные механизмы потенциала действия |
105 |
мембрану при том или ином потенциале (Vm), поскольку:
Однако, прежде необходимо решить две проблемы. Во-первых, ток, протекающий через мембрану, приведет к изменению мембранного потенциала; это, в свою очередь, вызовет изменение проводимостей. Решение было найдено путем разработки метода, позволяющего быстро устанавливать заданный мембранный потенциал и поддерживать его на постоянном уровне, при этом измеряя величину и временной ход мембранного тока. Поскольку потенциал фиксирован в течение всего времени эксперимента, ток будет аккуратно отражать изменения проводимости мембраны. Вторая проблема состоит в том, чтобы разделить различные компоненты мембранного тока для того, чтобы измерить их индивидуальные характеристики. Для преодоления этого затруднения применялись различные методы, в том числе замена натрия на ион, не проникающий через мембрану, а позднее использовались селективные токсины и яды.
§ 2. Эксперимент с фиксацией потенциала
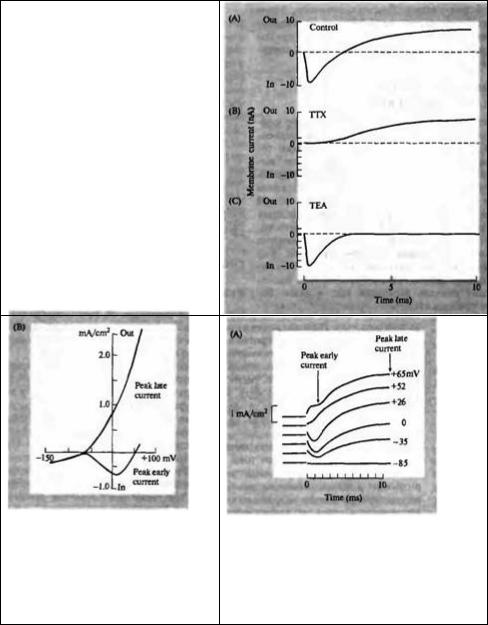
Метод фиксации потенциала (voltage clamp) был разработан Колем10, 11) и его коллегами, и усовершенствован Ходжкином, Хаксли и Катцем6). Все, что нам необходимо знать для того, чтобы понять сами эксперименты, это то, что метод позволяет практически мгновенно установить нужный уровень мембранного потенциала и зафиксировать его на этом уровне, измеряя при этом мембранный ток. На рис. 6.3А показан пример тока, протекающего через мембрану при скачкообразном изменении потенциала с уровня покоя (–65 мВ) на деполяризованный уровень (–9 мВ). Ток, возникающий в ответ на скачок потенциала, имеет три стадии: (1) короткий выброс выходящего тока, длящийся всего несколько миллисекунд, (2) ранний входящий ток и (3) поздний выходящий ток.
Емкость и ток утечки
Первоначальный кратковременный выброс тока представляет собой емкостной ток, обусловленный изменением заряда на мембране в результате изменения мембранного потенциала. Если усилитель обратной связи способен проводить большие токи, то емкостной ток длится очень недолго. На практике выброс емкостного тока длится около 20 мс, и за ним следует небольшой, но устойчивый выходящий ток.
Этот выходящий ток, протекающий через проводимости, активные при потенциале покоя, называется током утечки. Большей частью это ток ионов калия и хлора, который имеет линейную зависимость от величины смещения потенциала фиксации от потенциала покоя и наблюдается на всем протяжении скачка потенциала. Большую часть времени, однако, этот ток замаскирован другими, гораздо большими по величине ионными токами.
Токи ионов натрия и калия
Ходжкин и Хаксли показали, что вторая и третья стадии тока обусловлены сначала входом ионов натрия, а затем выходом ионов калия из клетки. Им также удалось выделить индивидуальные компоненты тока и рассчитать их величину и временной ход. Одним из удобных способов добиться этого послужило удаление из раствора большей части ионов натрия и замена их на ионы холина (которые не проходят через мембрану). Снизив содержание внеклеточного натрия, удалось добиться того, что натриевый равновесный потенциал сравнялся с деполяризованным мембранным потенциалом (–9 мВ на рис. 6.3А). Таким образом, суммарный ток натрия равнялся нулю. Остался только калиевый ток, который показан на рис. 6.3D. Натриевый ток (рис. 6.3С) был получен путем вычитания калиевого компонента из суммарного тока (рис. 6.3А).
Избирательные яды для натриевых и калиевых каналов
Со времен первоначальных опытов Ходжкина и Хаксли были найдены удобные фармакологические средства, позволяющие избирательно блокировать натриевые или калиевые токи. Особенно удобным средством блокирования натриевых токов оказался тетродотоксин (ТТХ) и его фармакологический спутник сакситоксин (STX) (глава 3). ТТХ представляет собой сильный яд, содержащийся в яичниках и других органах рыбы фугу. В своем обзоре Као 12) описывает захватывающую историю этого яда, начинающуюся с его

106 |
Раздел II. Передача информации в нервной системе |
Рис. 6.3. Мембранные токи в ответ на деполяризацию. (А) Общий ток. (В) Емкостной ток. (С) Ранний входящий ток. (D) Поздний выходящий ток.
Fig. б.З. Membrane Currents produced by depolarization. (A) Currents measured by a voltage clamp during a 56 mV depolarization of a squid axon membrane. The currents (lower trace) consist of a brief positive capacitative current an early transient phase of inward current and a late, maintained outward current. These are shown separately in В, С and D. The capacitative current (B) lasts for only a few microseconds (note the change in timescale). The small outward leak current is due to the movement of potassium and chloride. The early inward current (C) is due to sodium entry, the late outward current
(D) to potassium movement out of the fiber.
открытия китайским императором Шун Нунгом (2838-2698 до н. э). Император лично испробовал 365 различных веществ, составляя свою фармацевтическую коллекцию, и прожил на удивление долго. STX синтезируется морским планктоном и накапливается в питающихся планктоном моллюсках, таких как аляскинский Saxidomus. По силе действия этот яд сравним с ТТХ; употребление в пищу всего одного такого моллюска (сырым или в вареном виде) может привести к смертельному исходу.
Важное преимущество ТТХ для нейробиологических опытов заключается в высокой избирательности его действия. Мур, Нарахаши и их коллеги в своих экспериментах на аксоне кальмара показали, что ТТХ в концентрации всего 1 мкмоль или ниже избирательно блокирует потенциалзависимые натриевые каналы13). Изменение потенциала фиксации аксона, обработанного ТТХ, не приводит к возникновению входящего натриевого тока, но лишь выходящего калиевого (рис. 6.4А и В). Величина и временной ход калиевого тока не изменяются в присутствии ТТХ. Внутриклеточное введение ТТХ в аксон никак не действует на ток. Эффект STX ничем не отличается от эффекта ТТХ. Оба токсина связываются с одним и тем же посадочным местом, расположенным во внешнем устье канала, через которое проходят ионы натрия, и физически перекрывают им проход через мембрану 14).
Для калиевых каналов, участвующих в потенциале действия, также было найдено несколько блокирующих их вешеств. Так, в
Глава б. Ионные механизмы потенциала действия |
107 |
Рис. 6.4. Фармакологическое различение мембранных токов. (А) Отведение в нормальном растворе. (В) Блок натриевого тока тетродотоксином. (С) Блок калиевого тока TEA.
Fig. 6.4. Pharmacological Separation of Membrane Currents into sodium and potassium components. Membrane currents were produced by clamping of the membrane potential to 0 mV in a frog myelinated nerve. (A) Control record in normal bathing solution. (B) The addition of 300 nW tetrodotoxin (TTX) causes the sodium current to disappear while the potassium current remains. (C) The addition of tetraethylammonium (TEA) blocks the potassium current leaving the sodium current intact. (After Hille, 1970.)
Рис. 6.5. Потенциалзависимость раннего и |
indicated. The late potassium current increases as the |
позднего токов. (А) Токи в ответ на гипер- и |
depolarizing steps increase. The early sodium current first |
деполяризацию. (В) Зависимость амплитуды |
increases, then decreases with increasing depolarization; it is |
тока от величины пре-гиперполяризации и |
absent at +52 mV and reversed in sign at +65 mV. (B) Peak |
пре-деполяризации. |
currents plotted against the potential to which the membrane |
Fig. 6.5. Dependence of Early and Late Currents |
is stepped. Late outward current increases rapidly with |
on Potential. (A) Currents produced by voltage |
depolarization. Early inward current first increases in |
steps from a holding potential of -65 mV to a |
magnitude, then decreases, reversing to outward current at |
hyperpolanzed level (-85 mV) and to |
about +55 mV (the sodium equilibrium potential). (After |
successively increasing depolarized levels as |
Hodgkin, Huxley, and Katz, 1952.) |
аксоне кальмара и миелинизированном аксоне лягушки, как показали Армстронг, Хилле и их коллеги, тетраэтиламоний (TEA, в концентрациях выше 10 ммоль) блокирует потенциалзависимые калиевые каналы (см. рис. 6.4С) 15).
В аксоне кальмара TEA необходимо добавлять во внутриклеточный раствор, поскольку он блокирует внутреннее устье канала; в других препаратах, таких как перехват Ранвье лягушки, TEA эффективен и при внеклеточ-
108 |
Раздел II. Передача информации в нервной системе |
ной аппликации. Другие соединения, такие как 4-аминопиридин (4-АР) или диаминопиридин (DAP), блокируют калиевые каналы в миллимолярных концентрациях как снаружи, так и изнутри клетки.
Зависимость ионных токов от мембранного потенциала
Установив природу раннего и позднего компонента тока, Ходжкин и Хаксли исследовали зависимость натриевого и калиевого токов от мембранного потенциала. Токи, вызванные различными уровнями деполяризации от потенциала покоя -65 мВ, показаны на рис. 6.5А. Скачок потенциала на -85 мВ (нижняя кривая на рис. 6.5А) приводит лишь к возникновению небольшого входящего тока, как и следовало ожидать на основании свойств мембраны в состоянии покоя. Как уже было показано на рис. 6.3, каждый из небольших деполяризационных скачков производит сначала входящий ток, а затем более продолжительный выходящий. С увеличением величины деполяризационных скачков ранний ток уменьшается, при деполяризации на +52 мВ он равен нулю, а при еще более положительных значениях деполяризации меняет знак и становится выходящим.
Потенциалзависимость ранней и поздней составляющих тока показана на рис. 6.5В. Максимальная амплитуда раннего тока и амплитуда уровня плато позднего тока расположены по оси ординат, а потенциал фиксации, который устанавливался скачком с уровня потенциала покоя — по оси абсцисс. При гиперполяризующих скачках нет разделения на ранние и поздние токи; мембрана отвечает на скачок потенциала как простой резистор. Поздний ток также ведет себя как резистор, поскольку деполяризация активирует выходящий ток, однако по мере увеличения уровня деполяризации величина тока начинает значительно превосходить величину, ожидаемую на основании свойств мембраны в покое. Это объясняется активацией потенциалзависимой калиевой проводимости, пропускающей дополнительный ток. Поведение раннего тока значительно более сложно. Как уже было отмечено, он сначала возрастает, а затем убывает по мере увеличения деполяризации, становится равным нулю при потенциале +53 мВ, а затем и вовсе меняет знак. Потенциал реверсии расположен близко к равновесному потенциалу для натрия, чего и следовало ожидать для тока ионов натрия.
Интересной особенностью потенциалзависимости раннего тока является то, что в диапазоне потенциалов от —50 до +10 мВ ток растет с ростом деполяризации. Величина натриевого тока зависит от натриевой проводимости (gNa), а также от движущей силы для ионов натрия (Vm — ЕNа). Можно было бы предположить, что благодаря этому ток будет уменьшаться по мере приближения мембранного потенциала к натриевому равновесному потенциалу, т. е. произойдет уменьшение движущей силы. Однако этого не происходит по причине резкого увеличения натриевой проводимости с возрастанием деполяризации (см. рис. 6.7), которое перевешивает эффект снижения движущей силы. Таким образом, натриевый ток INa = gNa(Km - ENa) возрастает. Этот участок потенциалзависимости раннего тока называется «участком проводимости с отрицательным углом наклона».
Инактивация натриевого тока
Из опытов Ходжкина и Хаксли очевидно, что временной ход натриевого и калиевого токов весьма различен. Развитие калиевого тока замедлено по сравнению с натриевым, однако по достижении своего максимального уровня калиевый ток остается на нем довольно долго. Напротив, натриевый ток растет гораздо быстрее, но затем спадает до нуля, несмотря на то, что мембрана все еще деполяризована. Такой спад натриевого тока называется инактивацией.
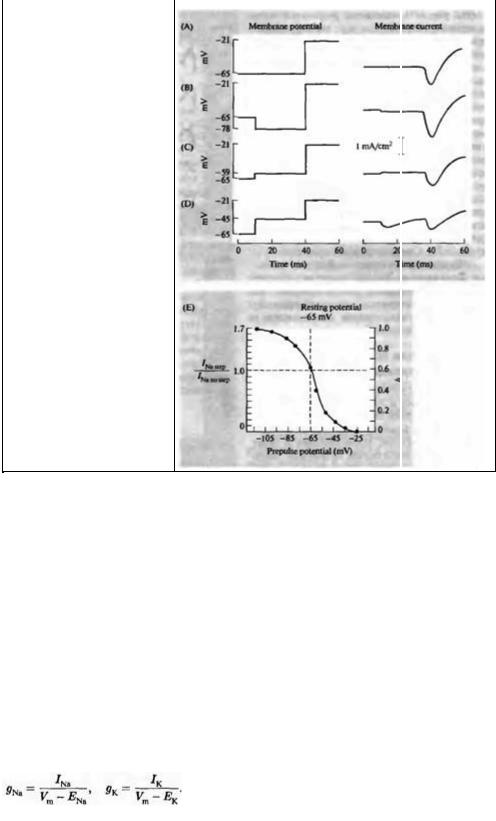
Ходжкин и Хаксли детально изучили процесс инактивации натриевого тока. В частности, они исследовали влияние гиперполяризующих и деполяризующих пре-импульсов на пиковую амплитуду натриевого тока, возникающего в ответ на последующую деполяризацию. Пример такого эксперимента показан на рис. 6.6. На рис. 6.6А мембранный потенциал скачкообразно переведен с -65 на -21 мВ, в результате чего возникает натриевый ток величиной приблизительно 1 мА/см2. Когда деполяризации предшествует гиперполяризация величиной —13 мВ, пиковое значение тока возрастает (рис. 6.6В). Деполяризующие пре-импульсы, напротив, снижают амплитуду натриевого тока (рис. 6.6С и D). Воздействие деполяризующих и гиперполяризующих пре-импульсов зависит от времени воздействия: короткие импульсы длиной в несколько миллисекунд неэффективны. В данном эксперименте длительность пре-
Глава б. Ионные механизмы потенциала действия |
109 |
|||
Рис. 6.6. |
Влияние |
мембранного |
|
|
потенциала на натриевый ток. (А) Ток |
|
|||
в ответ на деполяризацию. (В) |
|
|||
Увеличение |
амплитуды тока |
после |
|
|
пре--гиперполяризации. (С и D) |
|
|||
Предварительная |
деполяризация |
|
||
снижает амплитуду входящего тока. |
|
|||
(Е) Зависимость относительного |
|
|||
изменения амплитуды тока от пре- |
|
|||
гиперполяризации |
и |
пре- |
|
|
деполяризации. |
|
|
|
|
Fig. 6.6. Effect of Membrane Potential on Sodium Currents. (A) A depolarizing step from -65 to -21 mV produces inward sodium current followed by outward potassium current. (B) When the depolarizing step is preceded by a 30 ms hyperpolanzmg step, the sodium current is increased. Prior depolarizing steps (C and D) reduce the size of the inward current. (E) The fractional increase or reduction of the sodium current as a function of membrane potential during the preceding conditioning step. The maximum current with a hyperpolarizing step to -105 mV is about 1,7 times larger than the control value. A depolarizing step to -25 mV reduces the subsequent response to zero. Full range of the sodium current is scaled from zero to unity by the h ordinale.
импульсов была достаточной (30 мс) для достижения максимального эффекта.
Результаты представлены количественно на рис. 6.6Е в виде зависимости пикового натриевого тока от потенциала пре-импульса. Пиковый ток выражен в долях амплитуды контрольного тока. При деполяризующем пре-импульсе порядка —30 мВ натриевый ток был снижен до нуля, т. е. инактивация была полной. Гиперполяризующие пре-импульсы до -95 мВ и ниже вызвали увеличение натриевого тока на 70 %. Ходжкин и Хаксли выразили этот диапазон значений натриевого тока от нуля до максимума одним параметром (h), принимающим значение от нуля (полная инактивация) до 1 (отсутствие инактивации), как показано на правой оси ординат на рис. 6.6Е. В данных экспериментах инактивация при потенциале покоя была около 40 % максимального значения. Последующие эксперименты показали, что натриевые каналы всех нейронов демонстрируют ту или иную степень инактивации в покое.
Натриевая и калиевая проводимость как функция потенциала
Измерив таким образом величину и временной ход натриевого и калиевого токов как функции мембранного потенциала Vm и определив равновесные потенциалы ENa и Ек, Ходжкин и Хаксли получили возможность рассчитать величину и временной ход изменений натриевой н калиевой проводимостей, используя приведенные ранее соотношения·

110 |
Раздел II. Передача информации в нервной системе |
Рис. 6.7. Натриевая и калиевая проводимости. (А) |
Peak sodium conductance and steady-state potassium |
||||
Изменения проводимости в ответ на скачок |
|||||
потенциала. (В) Зависимость натриевой и калиевой |
conductance |
both |
increase |
with |
increasing |
проводимостей от потенциала во время скачка. |
depolarization. (B) Peak sodium conductance and |
||||
Fig. 6.7. Sodium and Potassium Conductances. (A) |
steady-state potassium conductance plotted against the |
||||
Conductance changes produced by voltage steps from — |
potential to which the membrane is stepped. Both |
||||
65 mV to the indicated potentials. |
increase steeply with depolarization between -20 and +10 |
||||
|
mV. (After Hodgkin and Huxley, 1952b.) |
|
|||
На рис. 6.7А показаны результаты пяти скачков потенциала. Как gNa, так и gк возрастают с увеличением деполяризации. Временной ход натриевой проводимости совпадает с натриевым током, однако ее потенциалзависимость существенно отличается от токовой (рис. 6.5). Проводимость равномерно нарастает с увеличением деполяризации, в то время как величина натриевого тока сначала возрастает, а затем убывает по мере увеличения деполяризационных скачков. Чем ближе мембранный потенциал во время деполяризации к натриевому равновесному потенциалу, тем меньше натриевый ток. В результате входящий ток уменьшается, несмотря на то, что проводимость растет. Зависимость максимальной проводимости для натрия и калия от мембранного потенциала показана на рис. 6.7В. Сходство кривых очевидно.
Обобщим вышесказанное. Результаты опытов Ходжкина и Хаксли показали, что деполяризация мембраны аксона вызывает три процесса: (1) активацию натриевой проводимости, (2) ее последующую инактивацию и (3) активацию калиевой проводимости.
Количественное описание натриевой и калиевой проводимостей
На основе полученных экспериментальных данных Ходжкин и Хаксли разработали математическую модель, способную точно описать временной ход изменений натриевой и калиевой проводимостей, происходящих в ответ на деполяризующий скачок потенциала. Если рассматривать калиевую проводимость, то логическим следствием изменения потенциала должно быть возникновение движущей силы, способной переместить один или несколько зарядов внутри потенциалзависимого калиевого канала, который в результате должен открыться. При условии, что речь идет об одном-единственном процессе, кинетика изменений калиевой проводимости должна описываться уравнением первого порядка, т. е. возрастание проводимости в ответ на деполяризацию должно быть экспоненциальным.
Вопреки таким рассуждениям, процесс увеличения калиевой проводимости начинается с задержкой и протекает по S-образной кривой (см. рис. 6.7А). Благодаря наличию этой задержки, а также тому, что увеличение калиевой проводимости возникает только в ответ на деполяризацию, но не гиперполяризацию, этот ионный канал получил название задержанного выпрямления (delayed rectifier). Ходжкин и Хаксли нашли способ для точного математического описания процесса увеличения калиевой проводимости, сделав допущение о том, что для для открытия канала необходима активация четырех процессов первого порядка (например, перемещение

Глава 6. Ионные механизмы потенциала действия |
111 |
четырех заряженных частиц внутри мембраны). Другими словами, S-образную кривую можно описать суммой четырех экспонент. Таким образом, калиевую проводимость можно представить как
где g K(max) — максимальная проводимость для данного скачка потенциала, а n — возрастающая экспоненциальная функция, принимающая значения от 0 до 1: n = 1 — е–t/τn. Зависимость gK(max) от потенциала показана на рис. 6.7. Временная константа экспоненты, τn, также зависит от потенциала: чем больше деполяризация, тем быстрее возрастает проводимость. При температуре 10° С τn принимает значения в диапазоне от 4 мс для небольших деполяризаций до 1 мс для деполяризации до нуля мембранного потенциала. Временной ход возрастания натриевой проводимости также имеет форму S-образной кривой, но описывается экспонентой, возведенной в третью степень. Напротив, спад натриевой проводимости в результате инактивации происходит по моноэкспоненциальной кривой. Для каждого конкретного скачка потенциала, общий временной ход изменений натриевой проводимости представляет собой результат наложения процессов активации и инактивации:
где gNa(max) — это максимальный уровень, которого натриевая проводимость достигла бы при отсутствии инактивации, a m = 1 — е—t/τn Процесс инактивации представлен не нарастающей, а спадающей экспонентой, представленной как h = е—t/τh. Как и в случае калиевой проводимости,
gNa(max) зависит от потенциала, также как и временные константы активации и инактивации. Временная константа активации натриевой проводимости τm гораздо короче калиевой, и при
10° С принимает значения от 0,6 мс (при значениях потенциала близких к потенциалу покоя) до 0,2 мс при нулевом мембранном потенциале. Временная константа инактивации τh близка по значению константе τn.
Реконструкция потенциала действия
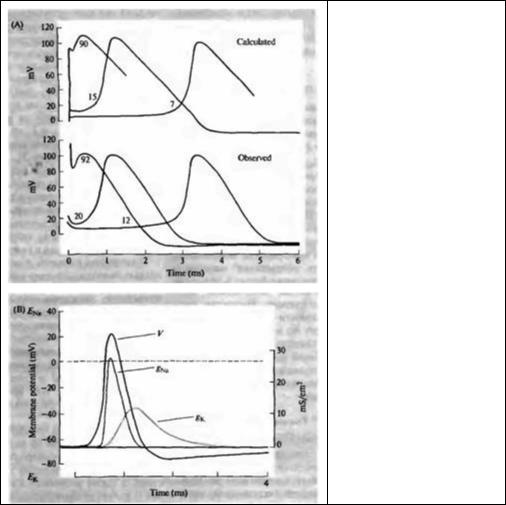
Эмпирически полученные формулы, отражающие зависимость натриевой и калиевой проводимостей от потенциала и от времени, позволили Ходжкину и Хаксли предсказать временной ход всего потенциала действия и происходящих при этом изменений проводимостей. Начав с деполяризующего скачка потенциала, лишь немного превышающего пороговое значение, они рассчитали все последующие изменения мембранного потенциала с шагом 0,01 мс. Так, они подсчитали, как в течение первых 0,01 мс после деполяризации до -45 мВ изменятся gNa и gк, каково будет увеличение INa и IK, и, наконец, каким будет в результате воздействие суммарного тока на Vm. Зная значение Vm в конце первого отрезка времени в 0,01 мс, они повторили те же вычисления для следующего промежутка, затем для следующего, и так далее, на протяжении всех фаз нарастания и спада потенциала действия (исключительно трудоемкая процедура, если учесть, что ни компьютеров, ни даже калькуляторов в те времена не было!).
Результаты расчетов с удивительной точностью повторили форму реального потенциала действия аксона кальмара. На рис. 6.8А приведены экспериментально измеренный и рассчитанный потенциалы действия при трех различных уровнях деполяризации. Для того, чтобы по достоинству оценить значимость этих достижений, нужно иметь в виду, что параметры, использовавшиеся для расчетов, были получены в совершенно неестественных условиях, при которых потенциал последовательно фиксировался на разных уровнях.
Схема механизма генерации потенциала действия представлена на рис. 6.8В, где показаны величина и временной ход рассчитанного распространяющегося потенциала действия, а также расчетные изменения натриевой и калиевой проводимостей.
Порог и рефрактерный период
Ходжкин и Хаксли не только описали потенциал действия, но объяснили многие другие свойства возбудимых аксонов, например, порог и рефрактерный период. Со временем выяснилось, что описанные ими закономерности применимы и ко многим другим возбудимым тканям.
Можно ли на основе этих закономерностей предсказать величину порогового мембранного потенциала, при котором запускается механизм потенциала действия? Не означает ли дискретность такого понятия, как порог, того, что ионная проводимость тоже должна быть дискретной (не непрерывной) функцией? Для того, чтобы понять данный фено-
112 |
Раздел II. Передача информации в нервной системе |
Рис. 6.8. Реконструкция потенциала действия. (А) Сравнение рассчитанного потенциала действия с реально измеренным. (В) Соотношение между потенциалом действия и изменениями натриевой и калиевой проводимостей.
Fig. 6.8. Reconstruction of the Action Potential. (A) Calculated action potentials produced by brief depolarizations of three different amplitudes (upper panel) are compared with those recorded under the same conditions (lower panel). (B) Relation between conductance changes
and gк) and the action potential (V), calculated for a propagated action potential in a squid axon. (After Hodgkin and Huxley, 1952d.)
мен, представим себе ситуацию, при которой потенциал мембраны увеличивается вплоть до достижения порогового значения, а затем ток отключается. Поскольку мембрана деполяризована, выходящий ток будет превышать величину тока в покое, протекающего через калиевые каналы (и каналы утечки). Некоторое количество натриевых каналов также будет активировано, поэтому натриевый ток возрастет. При пороговом значении мембранного потенциала входящий и выходящий токи также уравновешивают друг друга, как и в состоянии покоя. Однако, есть одно важное отличие от покоя: равновесие это неустойчиво. Если в клетку войдет хотя бы один ион натрия, то возрастет деполяризация, возрастет и натриевая проводимость, в результате чего в клетку войдет еще большее количество натрия. Выходящий ток в такой ситуации уже не может компенсировать входа натрия, поэтому возникает регенеративный процесс. С другой стороны, если теперь из клетки выйдет один ион калия, то деполяризация уменьшится, снизится и натриевая проводимость, в результате чего избыточный выходящий ток приведет к гиперполяризации. По достижении уровня потенциала покоя калиевый ток снизится до величины противоположного ему натриевого тока.