

Nikolls_-_Ot_neyrona_k_mozgu
.pdfГлава 8. Свойства и функции нейроглиальных клеток |
153 |
Рис. 8.10. Нейроны мигрируют вдоль радиальной глии в процессе развития. (А) Срез затылочной доли развивающейся коры плода обезьяны в середине беременности. Радиальные глиальные волокна идут от вентрикулярной зоны (внизу) к поверхности развивающейся коры (вверху). (В) Мигрирующая клетка
(1) имеет ведущий отросток, располагающийся на радиальной глиальной клетке и использующей его в качестве проводника. Клетка 2, которая продвинулась дальше, оставляет за собой отросток, также прикрепленный к глиальной клетке. Клетка 3 только начинает образовывать отросток, прежде чем начать свою миграцию вдоль глиальной клетки. (С) Миграция гиппокампального нейрона вдоль радиального глиального волокна (GF) in vitro. С течением времени лидирующий отросток (LP) продвигается все выше и выше, а за ним следует тело нейрона. Время в минутах указано внизу.
Fig. 8.10. Neurons Migrating along Radial Glia during development. (A) Camera lucida drawing of the occipital lobe of developing cortex of a monkey fetus at mid-gestation. Radial glial fibers run from the ventricular zone below to the surface of the developing cortex above. (B) Three-dimensional reconstruction of migrating neurons. The migrating cell (1) has a voluminous leading process that follows the radial glia, using it as guideline. Cell 2, which has migrated farther, retains a process still connected to the radial glia. Cell 3 is beginning to send a process along the radial glia before migrating. (C) Migration of a hippocampal neuron along a radial glial fiber (GF) in vitro. As time progresses, the leading process (LP) moves farther up, with the neuronal cell body following. Times indicated at the bottom represent real time, in minutes, taken from video photography. (A and В after Rakic, 1988; С from Hatten, 1990.)
на рис. 8.10. Мигрирующие нейроны распознают на поверхности глиальных клеток поверхностные молекулы, специфические к их нейрональному типу. Например, радиальные глиальные клетки из гиппокампа могут направлять миграцию нейронов гиппокампа, но не неокортекса.
Роль микроглиальных клеток в репарации и регенерации в ЦНС
Астроциты, микроглия и шванновские клетки реагируют на нейрональное повреждение репликацией 6· 7). Они участвуют в удалении продуктов распада и в рубцевании. На первом этапе резидентные микроглиальные клетки и макрофаги, которые проникают в
поврежденный участок ЦНС из крови, делятся и удаляют продукты распада умирающих клеток 47).
Мюллер и его сотрудники исследовали роль микроглиальных клеток в регенерации ЦНС у пиявки 48). (Кстати, именно у пиявки микроглиальные клетки были впервые описаны и получили свое название 49). В нормальных условиях микроглиальные клетки равномерно распределены в ганглиях и пучках аксонов, которые их соединяют (рис. 8.11). Сразу после повреждения ЦНС микроглиальные клетки мигрируют к месту повреждения со скоростью около 300 мкм/час, аккумулируются на этом месте и фагоцитируют поврежденную ткань. Гибридизация in situ и иммунохимические реакции показали, что микроглиальные клетки продуцируют в месте повреждения ламинин 50) (глава 24) — молекулу

154 |
Раздел II. Передача информации в нервной системе |
Рис. 8.11. Миграция |
микроглиальных клеток к |
|
|
|
||||
участку повреждения в |
ЦНС. (А) Микроглия в |
|
|
|
||||
ЦНС пиявки, окрашенная флуоресцентным |
|
|
|
|||||
красителем (Hoechst 33342). Пучок аксонов, |
|
|
|
|||||
связывающих ганглии, был сдавлен 5 минут |
|
|
|
|||||
назад. Участок сдавления отмечен пунктирной |
|
|
|
|||||
линией. Микроглиальные клетки все еще |
|
|
|
|||||
|
|
|||||||
равномерно распределены. (В) Через три часа |
|
The bundle of axons linking ganglia had been crushed 5 minutes earlier. The |
|
|||||
после повреждения Микроглиальные |
клетки |
|
|
|
||||
extent of the crush is indicated by the dotted Line.The nuclei of microglial |
||||||||
собираются на месте сдавления и начинают |
|
cells were still evenly distributed at this time. (B) Three hours after the |
||||||
производить ламинин (С) Скорость и |
|
injury, microglial cells had accumulated at the crush site. There they |
||||||
расстояние, |
пройденное микроглиальными |
|
produced the growth-promoting molecule laminin. (C) Velocities and |
|||||
клетками |
по |
направлению |
к |
месту |
|
distances traveled by microglial cells as they moved toward a lesion in leech |
||
повреждения. |
|
|
|
|
|
CNS. Microglial cells were tracked by video-microscopy at 10-min intervals |
||
|
|
|
|
|
|
|
||
Fig. 8.11. Migration of Microglial Cells in Injured |
|
in injured leech preparations. In uninjured preparations, microglial cells |
||||||
CNS. (A) Microglia in the Leech CNS were stained |
|
make only short random movements. (A and В after Chen et al., 2000, |
||||||
with a fluorescent nuclear dye (Hoechst 33342). |
|
micrographs kindly provided by K.J.Muller; С after McGlade-McCulloh et |
||||||
|
al., 1989.) |
|||||||
|
|
|
|
|
|
|
||
экстраклеточного матрикса, которая способствует росту нейритов в культуре и in vivo, как у пиявки, так и у позвоночных.
Шванновские клетки как пути роста в периферических нервах
Управление ростом аксонов нейронов шванновскими клетками было исследовано в экспериментах Thompson с коллегами на модели концевой пластинки — синапса между двигательным нервным окончанием и скелетной мышцей 51) - 53) (глава 24; рис. 8.12А). Икроножная мышца у взрослых крыс была частично денервирована. Было обнаружено, что неповрежденные аксоны разветвляются и устанавливают контакты с денервированными волокнами. В подобных условиях один аксон в состоянии установить контакты с количеством волокон, в 5 раз превышающим таковое в нормальных условиях. Рост аксонов был визуализирован с помощью антител к нейрофиламентам (рис. 8.12В). Шванновские клетки были окрашены другим специфическим антителом (рис. 8.12В). Прямое наблюдение выявило, что первыми начинают расти шванновские клетки денервированных волокон, направляя отростки к интактным аксонам (рис. 8.12А, с). Только после этого аксоны дают отросток, который следует по пути, сформированному отростком шванновской клетки. Интересно, что имплантация шванновской клетки рядом с неповрежденным аксоном также стимулировала спраутинг аксона, даже в отсутствии денервированного волокна.
В серии других экспериментов периферический нерв был полностью перерезан. Как и предполагалось, после короткой задержки аксоны начали расти из проксимального конца нерва. Однако, как и в предыдущем эксперименте, первым этапом был рост шванновских клеток, по отросткам которых аксоны могли расти к своим мишеням.
Замечание
Хотя эксперименты на нервно-мышечном синапсе ясно продемонстрировали роль шванновских клеток в направлении роста нейритов во время регенерации, следует быть осторожным при обобщении этого правила. Например, в ЦНС пиявки поврежденные аксоны могут расти и восстанавливать свои исходные связи даже после уничтожения всех

Глава 8. Свойства и функции нейроглиальных клеток |
155 |
Рис. 8.12. |
Шванновские |
клетки |
— |
проводники |
аксонов, |
растущих |
к |
денервированным концевым пластинкам на скелетных мышечных волокнах. (А) Схема событий, происходящих при частичной денервации мышцы крысы: (а) контроль; (b) аксон, иннервирующий волокно справа, был перерезан, что привело к дегенерации нервного окончания; (с) шванновская клетка, остающаяся на денервированном мышечном волокне, прорастает, и один из отростков достигает нервного окончания на близлежащем мышечном волокне; (d) от аксона отрастает новая ветка, которая растет вдоль отростка шванновскои клетки к денервированной концевой пластинке и устанавливает с ней синаптический контакт. (В) Рост вновь образованной ветки аксона (окрашенной антителами к нейрофиламентам) к денервированн му синапсу вдоль отростка шванновской клетки (окрашенной специфическими к шванновским клеткам антителами 4E2). Спустя три дня после частичной денервации нервная ветка проросла от неповрежденного аксона к денервированной мышце, следуя вдоль отростка шванновской клетки, который образовался ранее.
Fig. 8.12. Role of Schwann Cells in Guiding Axons to denervated motor end plates of skeletal muscle fibers. (A) Schematic representation of the effects of partial denervation of a rat muscle, (a) At normal nerve-muscle synapses, an axon and its Schwann cell are closely apposed. (b) The axon innervating the right-hand muscle fiber is cut leading to degeneration of the nerve terminal, (c) In response to this denervation, the terminal Schwann cells on the denervated muscle fiber grow processes, one of which reaches the nerve terminal at the adjoining muscle fiber, (d) An axonal sprout is induced from the uninjured nerve terminal. It grows along the Schwann cell process to the denervated end plate, which it rein-nervates.
(B) Growth of axonal sprouts (labeled with antineurofilament antibody) to denervated synapses along processes extended by Schwann cells (labeled with a monoclonal antibody, 4E2, which is specific for Schwann cell bodies and processes). Three days after partial denervation, a neurofilament-labeled nerve sprout has grown from the innervated junction to a denervated junction by following the Schwann cell process that had grown earlier. The innervated and denervated motor end plates were identified by the patterns of staining of axons and Schwann cells. (After Son and Thompson, 1995; micrographs kindly provided by W. Thompson.)
156 |
Раздел II. Передача информации в нервной системе |
окружающих их глиальных клеток. Большое количество синапсов устанавливается в ЦНС на эмбриональном этапе развития, когда количество глиальных клеток очень мало. Более того, даже при полном отсутствии глиальных клеток вполне нормальные синапсы формируются в культуральных условиях. Таким образом, маловероятно, что «синапсы не могут ни формироваться, ни функционировать в отсутствии глиальных клеток» 54).
§ 3. Эффекты нейрональной активности на глиальные клетки
Накопление калия во внеклеточном пространстве
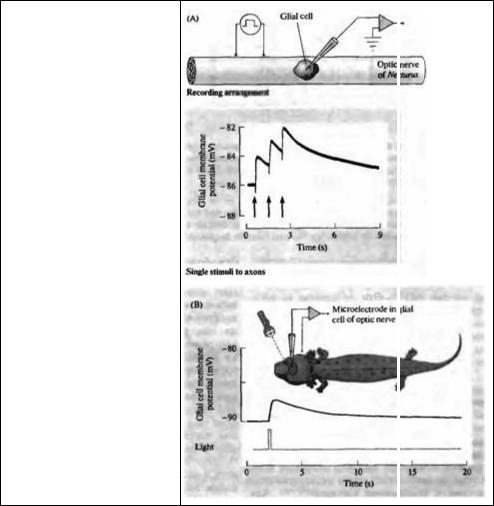
Деполяризация глиальной клетки во время активности нейрона проиллюстрирована на рис. 8.13. Регистрация производилась от глиальной клетки в зрительном нерве молодого тритона (Necturus). Потенциалы действия, запускаемые в нервных волокнах электрической стимуляцией или вспышками света, проходят мимо глиальной клетки и приводят к ее деполяризации, которая суммируется при прохождении нескольких импульсов 55). В коре млекопитающих глиальные клетки также деполяризуются, причем деполяризация тем больше, чем больше и чаше активировано рядом расположенных нейронов при стимуляции нервных трактов, периферических нервов, поверхности коры и чувствительных входов 56).
Причиной деполяризации глии является выход калия из аксонов. При накоплении калия во внеклеточном пространстве соотношение [K]0/[K]i меняется, что и приводит к изменению мембранного потенциала глиальных клеток. Калий-чувствительные электроды были использованы для измерения накопления калия во внеклеточном пространстве во время нейрональной активности 57).
Изменение мембранного потенциала глиальных клеток косвенно отражает уровень проходящих около них нервных импульсов. Калиевая сигнализация между нейронами и глиальными клетками отличается от специфической синаптической активности. Синаптическое взаимодействие ограничивается специализированными участками на телах и дендритах нервных клеток, и может быть как возбуждающим, так и тормозящим.
В противоположность этому, сигнализация калием не ограничивается структурами, содержащими рецепторы, и срабатывает независимо от того, что послужило причиной увеличения внеклеточной концентрации калия. В отличие от глиальных клеток, нейроны деполяризуются при повышении внеклеточного К в меньшей степени, поскольку зависимость их мембранного потенциала от наружного К отклоняется от уравнения Нернста в физиологическом диапазоне концентраций (глава 5).
Прохождение токов и движение калия через глиальные клетки
Между участками клетки, имеющими разные потенциалы, протекают токи. Нервные клетки используют этот закон для проведения импульса: токи проходят между невозбужденным участком аксона и той частью, по которой в данный момент проводится потенциал действия. Поскольку глиальные клетки связаны друг с другом контактами с низким сопротивлением, по проводящим свойствам они схожи с одной удлиненной клеткой. Поэтому деполяризация нескольких глиальных клеток при повышении концентрации внеклеточного калия в их окружении приводит к переносу зарядов от клеток, которые не подверглись воздействию. По тому же принципу удлиненная мюллеровская клетка, которая проходит через всю толщину сетчатки, генерирует ток при повышении концентрации калия лишь на небольшой части ее поверхности (рис. 8.6 и 8.14). Входящий ток в участке повышения [К]0, переносимый ионами калия, распространяется к другим областям глиальной клетки и через щелевые контакты — к другим глиальным клеткам. Токи, генерируемые глиальными клетками, вносят вклад в сигналы, регистрируемые внеклеточными электродами от сетчатки (электроретинограмма) или с поверхности головы (электроэнцефалограмма). Такого рода исследования дают весьма ценную информацию при клинической диагностике заболеваний.
Глия как буфер экстраклеточной концентрации калия
Одним из очевидных свойств глии является разделение и группировка нейрональных отростков. В результате этого концентрации
Глава 8. Свойства и функции нейроглиальных клеток |
157 |
Рис 8.13. Влияние нейрональной активности на мембранный потенциал глиальных клеток в оптическом нерве тритона. (А) Одновременное возбуждение нервных волокон, вызываемое электрической стимуляцией, приводит к деполяризации глиальной клетки. Амплитуда деполяризации зависит от количества активированных аксонов и частоты стимуляции. (В) Вспышка света длительностью 0,1 с вызывает деполяризацию глиальной клетки в оптическом нерве in vivo.
Fig. 8. 13. Effect of Action Potentials on dial Cells in mud puppy optic nerve. (A) Synchronous impulses evoked by electrical stimulation of nerve fibers cause glial cells to become depolarized. The amplitude of the potentials depends on the number of axons activated and on the frequency of stimulation. (B) Illumination of the eye with a 0,1 s flash of light causes depolarization of a glial cell in the optic nerve of an anesthetized mud puppy, with intact circulation. Lower trace monitors light stimulus. (After Orkand, Nicholls, and Kuffler, 1966.)
калия может увеличиваться возле некоторых нейронов, в то время как другие оказываются защищенными глиальными клетками. Это легло в основу привлекательной гипотезы о том, что глиальные клетки регулируют концентрацию калия во внеклеточном пространстве и служат «пространственным буфером»16-58'. В соответствии с этой гипотезой глиальные клетки опосредуют захват ионов калия из внеклеточного пространства, поддерживая постоянство окружающей среды. Поскольку глиальные клетки связаны друг с другом, калий может входить в одном участке и выходить в другом, как было описано выше (рис. 8.14). Очевидно, что в результате повышения концентрации калия, он движется через глиальные клетки. Однако, довольно трудно оценить, сколько калия движется по глиальным клеткам и в какой степени это движение важно для уменьшения внеклеточной концентрации калия. Были сделаны попытки оценить роль этого процесса теоретически, на основании предположений о
геометрии, проводимости, диффузии и активном транспорте калия в нейронах и глиальных клетках 59).
Эффекты медиаторов на глиальные клетки
Такие медиаторы как ГАМК, глутамат, глицин и ацетилхолин, действуют на мембраны глиальных клеток, вызывая деполяризую-

158 |
Раздел П. Передача информации в нервной системе |
|
|
|
|
Рис. 8.14. Калиевые каналы в глиальных клетках. Глиальные клетки соединены друг с другом щелевыми контактами. Выделенный активными аксонами калий деполяризует близлежащие глиальные клетки и входит в них, вызывая ток и выходящее движение ионов калия по всей глиальной сети. Таким образом, глиальные клетки могут выполнять роль пространственного буфера внеклеточного калия.
Fig. 8.14. Potassium Currents in dial Cells. The glial cells in the diagram are linked by gap junctions. Potassium released by active axons in one region depolarizes the glial cell and enters, causing current flow and outward movement of potassium elsewhere in the glial tissue. The concept of spatial buffering of potassium has been postulated as a mechanism for influencing neuronal function by glial cells.
щие или гиперполяризующие ответы 25, 60) - 62). Активация ГАМКА рецепторов при воздействии ГАМК на мюллеровские клетки показана на рис. 8.15 63). Эти рецепторы во многом, хотя и не во всем, схожи с нейрональными ГАМКА рецепторами. Физиологическая роль рецепторов к нейромедиаторам на глиальных клетках пока еше не определена.
Вместе с тем, глиальные клетки играют важную роль в захвате медиаторов в ЦНС, как при физиологических, так и при патологических условиях. Уменьшение внеклеточной концентрации выделяемых в синапсах медиаторов (глутамата, норадреналина, глицина) происходит отчасти в результате простой диффузии, однако, основную роль в этом процессе играет захват медиаторов нейронами и глиальными клетками 64) - 66). Как и в нейронах, транспорт глутамата в глиальные клетки сочетается со входом натрия по его электрохимическому градиенту (глава 4). Если механизм удаления не работает, внеклеточная концентрация глутамата достигает очень высокого уровня, приводя к избыточной активации NMDA рецепторов, массивному входу кальция и клеточной смерти. Количественные расчеты показывают, что система транспорта, сосредоточенная в глиальных клетках, играет ключевую роль в предотвращении избыточного накопления глутамата во внеклеточном пространстве. Было также показано, что у трансгенной мыши, у которой в астроцитах отсутствует глутаматный транспортер GLT-1, развивается эпилепсия и повышенная чувствительность к конвульсантам 25).
Освобождение медиаторов глиальными клетками
При значительной деполяризации глиальных клеток под воздействием повышенного внеклеточного калия или глутамата, а также при повышении внутриклеточной концентрации ионов натрия, глиальные клетки начинают выделять глутамат во внеклеточное пространство 67). В основе этого процесса лежит механизм обратного транспорта, описанный в главе 4. На рис. 8.16 изображен эксперимент, показывающий электрический ток, который возникает при освобождении глутамата глиальными клетками. Процесс обратного транспорта может в значительной степени усугубить последствия повреждения мозга. Поврежденные и погибающие нервные клетки освобождают глутамат и К+ и таким образом деполяризуют глиальные клетки (и другие нейроны), которые в свою очередь освобождают еще больше глутамата.
Одним из лучших примеров освобождению медиатора глиальными клетками является регенерация в периферической нервной системе. В денервированных двигательных концевых пластинках шванновские клетки занимают места, освобождаемые двигательными нервными
окончаниями, и выделяют ацетилхолин, приводя к генерации миниатюрных потенциалов в мышце 68).
Кальциевые волны в глиальных клетках
У глиальных клеток, растущих в культуре, а также и in situ, можно наблюдать фено-

Глава 8. Свойства и функции нейроглиальных клеток |
159 |
|
|
Рис. 8.15. Ответы глиальных клеток на ГАМК (А) Ток, вызываемый ГАМК (GАВА) в мюллеровской глиальной клетке сетчатки. Концентрационная зависимость ответов, вызываемых ГАМК (справа). (В) Ответы, вызываемые ГАМК, блокируются бикукуллином — антагонистом ГАМКA рецепторов.
Fig. 8.15. Responses of dial Cells to GA6A. Responses of Muller glial cells in skate retina to GABA. (A) Current induced by GABA (30 µΜ) in a glial cell voltage-clamped at 0 mV. The dose-response relation for the peak of the GABA current is shown on the right (error bars indicate standard error of the mean). (B) The effect of GABA was blocked by bicuculline. a GABAA antagonist. (After Qian et al., 1996.)
мен сетевого увеличения внутриклеточного кальция, выделяемого из внутриклеточных депо (рис. 8.17). Используя флуоресцентные зонды, можно наблюдать эти осцилляторные волны (или «кальциевые спайки»), проходящие через щелевые контакты между глиальными клетками 69, 70). Кальциевые волны могут запускаться деполяризацией, медиаторами, механической стимуляцией. Они напоминают кальциевые волны в нейрональных сетях 71) (глава 23). Newman с коллегами показали, что распространяющиеся кальциевые волны могут запускать освобождение глутамата глиальными клетками в сетчатке, что, в свою очередь, может влиять на паттерны нейрональной активности 70). Физиологическая роль этих волн пока неизвестна.
Перенос метаболитов от глиальных клеток к нейронам
Исследования на пчелах и позвоночных показали, что глиальные клетки могут участвовать в доставке к нейронам питательных веществ 72) - 75). В агрегатах фоторецепторов и мюллеровских клеток, изолированных из сетчатки морской свинки, было показано, что молочная кислота (лактат), выработанная глиальными клетками из гликогена, накапливается во внеклеточной жидкости. В условиях повышенной нейрональной активности лактат переносится от глиальных клеток к фоторецептивным нейронам. Наличие такого механизма в культуральных условиях предполагает, что и в интактной ЦНС глиальные клетки могут участвовать в снабжении ней-

160 |
Раздел II. Передача информоции в нервной системе |
|
|
|
|
Рис. 8.16. |
|
Освобождение |
глутамата |
|
глиальными |
|
клетками. Освобождение |
|
|
глутамата |
в |
результате |
реверсии |
|
транспортера |
|
глутамата |
в |
|
мюллеровских клетках. (А) Вызываемое |
|||
|
деполяризацией освобождение глутамата из |
|||
мюл леровской клетки (справа) определяется по вызываемому освобождающимся глутаматом току в расположенной рядом клетке Пуркинье (слева). Клетка Пуркинье используется в качестве высокочувствительного детектора глутамата. (В) Деполяризация мюллеровской клетки от —60 до +20 мВ вызывает в близлежащей клетке Пуркинье входящий ток, опосредованный глута матными рецепторами. Ответ в клетке Пуркинье исчезает, если клетка Пуркинье удаляется от мюллеровской клетки (С), а также при удалении калия из внеклеточного раствора (D). В растворе, содержащем 0 ммоль К–, обратный транспорт глутамата из мюллеровской клетки блокируется.
Fig. 8.16. Release of Glutamate by dial Cells. Release of glutamate generated by reversal of the glutamate uptake carrier in a Muller cell.
(A) Depolarization--induced release of glutamate from a Muller cell (right) is monitored by recording glutamate-elicited currents from an adiacent Purkinie cell (left).
The Purkinje cell acts as a detector with high sensitivity and time resolution. (B) Depolarization of the Millier cell from -60 to +20 mV (top trace) elicits an inward current in the nearby Purkinje cell. The Purkinje cell current is generated by activation of its glutamate receptors. The response to glutamate disappears when the Purkinje cell is moved away from the Miiller cell (C) or when extracellular К > is omitted (D). In fluid containing 0 mAf К · , reverse glutamate transport by the Millier cell is blocked. (After Billups and Attwell, 1996.)
ронов питательными веществами при повышенной активности или же при недостатке кислорода.
Эффекты глиальных клеток на нейрональную сигнализацию
Описанные в предыдущих разделах эксперименты показывают важную роль глиальных клеток в развитии, регенерации и формировании миелина. На сегодняшний день довольно хорошо изучены также ионные каналы в мембранах глиальных клеток и роль глии в регуляции внеклеточной среды. Многие предположения о роли глиальных клеток были сделаны на основании экспериментов, проведенных в культуральных условиях, что не означает, что они верны in vivo. Вдобавок свойства глиальных клеток сильно варьируют в различных областях ЦНС. Например, оптический нерв, хотя и является распространенным препаратом для исследования глии 76), в то же время не является наиболее репрезентативным препаратом для исследования глии в целом, поскольку в нем нет синапсов. До сих пор нет достаточных доказательств тому, что глиальные клетки играют важную роль в динамических процессах, таких, например, как рефлекс на растяжение, или в ответах комплексных клеток зрительной коры в нормальных физиологических условиях (глава 20).
§ 4. Глиальные клетки и гематоэнцефалический барьер
Близкое расположение глиальных клеток, капилляров и нейронов в мозге предполагает.
Глава 8. Свойства и функции нейроглиальных клеток |
161 |
||
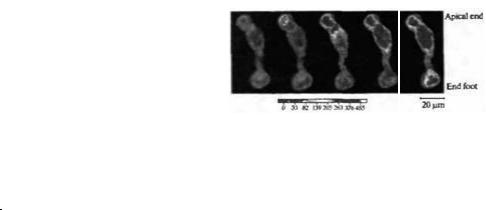
Рис. 8.17. Кальциевые |
волны в мюллеровских |
|
|
клетках саламандры. Кальциевые волны, вызванные |
|
|
|
добавлением рианодина в отсутствие внеклеточного |
|
|
|
кальция, начинаются с апикального конца клетки и |
|
|
|
распространяются по направлению к концевой ножке. |
|
|
|
Внутриклеточная концентрация кальция определялась |
|
|
|
с помощью кальций-чувствительной метки fura-2 с |
|
|
|
интервалом 7 секунд. Кальциевые волны могут также |
|
|
|
распространяться между глиальными клетками через |
|
|
|
щелевые контакты. Схожие волны могут запускаться |
|
|
|
физиологическими стимулами. Эти волны также |
The |
intracellular Ca2+ concentration was imaged using the Са2+ - |
|
напоминают волны в |
ней рональных клетках на |
indicator dye fura-2. Images were obtained at 7 s intervals. With glial |
|
ранних этапах развития. |
|
||
|
cells that are coupled by gap junctions, the calcium waves can spread |
||
|
|
||
Fig. 8.17. Calcium Wave in a Dissociated Salamander |
from cell to cell in a continuous progression. Similar waves are |
||
Miiller Cell. The Ca2+ wave, elicited by addition of 100 |
initiated by physiological stimuli. They resemble those seen in |
||
nM ryanodine in the absence of extracellular Ca2+, begins |
neurons during development. (After Keirstead and Miller, 1997; |
||
at the apical end of the cell and travels toward the cell end |
image kindly provided by S. Keirstead.) |
||
foot. |
|
|
|
что глиальные клетки могут быть частью гематоэнцефалического барьера. Гематоэнцефалический барьер располагается в местах соединения между специализированными эндотелиальными клетками, выстилающими кровеносные капилляры в мозге 77). Роль глиальных клеток была показана при вырашивании эндотелиальных клеток и астроцитов в культуральных условиях 78, 79). При выращивании без астроцитов эндотелиальные клетки редко соединяются друг с другом. Однако, в присутствии астроцитов происходит формирование полноценных плотных контактов, таких как in vivo. Эти контакты, которые полностью перекрывают межклеточное пространство между эндотелиальными клетками, составляют основу для непроницаемости мозговых капилляров. Для того, чтобы проникнуть из крови в мозг, молекулы должны пройти через эндотелиальные клетки, а не между ними. В свою очередь, наличие в культуре эндотелиальных клеток из капилляров мозга вызывает появление в астроцитах скоплений мембранных частиц. Взаимодействия между астроцитами и эндотелиальными клетками мозговых капилляров носят специфический характер. Фибробласты или эндотелиальные клетки из периферических сосудов не дают схожего результата. В настоящее время исследуются способы, дающие возможность вызвать разобщение эндотелиальных клеток, с тем чтобы увеличить проницаемость капилляров. Это позволило бы обойти гематоэнцефалический барьер (см. также следующий раздел) и обеспечить доставку в мозг тех фармакологических препаратов, которые в нормальных условиях через него не проходят 80).
Предположение о роли астроцитов в кровоснабжении мозга
Три факта свидетельствуют о возможной роли астроцитов в мозге млекопитающих. Вопервых, они окружают капилляры мозга своими концевыми отростками (именно это свойство привело Гольджи и многих других к предположению о том, что они занимаются доставкой веществ к нейронам). Во-вторых, активность в определенном участке мозга вызывает значительное увеличение кровоснабжения через этот участок, что отчетливо видно при использовании позитронной томографии (positron emission tomography, PET), магнитного резонанса (magnetic resonance imaging, MRI) и оптической регистрации. В-третьих, глиальные клетки отслеживают общий уровень активности в своем окружении. Паульсон и Ньюман 86) предложили интересную гипотезу о том, что концевые отростки деполяризованных астроцитов могут влиять на капилляры и вызывать местное расширение кровеносных сосудов. Посредством глиальной сигнализации активные нейроны могли бы получать таким образом дополнительное количество кислорода и глюкозы. Относительно механизмов можно предполагать, что освобождение калия, протонов или закиси азота (NO), освобождаемых из концевых отростков астроцитов, может влиять на эндотелиальные клетки капилляров. Предположение Paulson и Newman сродни исходному предположению Гольджи, однако в их гипотезе сигналы имеют обратное направление: не глиальные клетки переносят питательные вещества через свою цитоплазму из крови к нейронам.

162 |
Раздел II. Передача информации в нервной системе |
а нейрональная активность вызывает локальное расширение кровеносных сосудов и увеличение кровоснабжения именно там, где это необходимо.
§ 5. Глиальные клетки и иммунные ответы в ЦНС
До совсем недавних пор считалось, что ткань центральной нервной системы находится вне контроля иммунной системы. Наличие гематоэнцефалического барьера, отсутствие лимфатической системы и сравнительная легкость, с которой приживаются участки мозга при пересадке, — все это предполагает отсутствие иммунного ответа на чужеродные антигены. В результате функции ЦНС не нарушаются при массивных аллергических реакциях, вызываемых, например, укусом пчелы. Однако, астроциты в культуре и in situ взаимодействуют с Т-лимфоцитами, чья активность или стимулирует, или подавляет. Получены доказательства в пользу того, что микроглия и активированные Т-лимфоциты могут входить в мозг и вызывать острое воспаление в мозговой ткани 87, 88). Роль глии во взаимодействии между нервной и иммунной системами остается актуальной и еще далекой от решения проблемой.
Выводы
·Глиальные клетки в мозге и шванновские клетки на периферии создают окружение нейронов.
·Олигодендроциты имеют короткие отростки и миелинизируют крупные аксоны.
·Астроциты окружают мозговые капилляры.
·шванновские клетки миелинизируют периферические аксоны и продуцируют трофические молекулы.
·Микроглиальные клетки удаляют продукты распада после повреждения нервной системы и вовлечены в воспалительные ответы в нервной системе.
·Глиальные клетки имеют более отрицательный потенциал, чем нейроны, и не генерируют потенциалы действия.
·Глиальные клетки электрически связаны друг с другом, но не с нейронами.
·Мембраны глиальных клеток содержат натриевые, калиевые, кальциевые ионные каналы, а также рецепторы, насосы и транспортеры.
·Глиальные клетки играют роль в развитии нервной системы и в поддержании гомеостаза среды, окружающей нейроны.
Рекомендуемая литература Обзоры
оKettenmann, H., and Ransom, В. R. (eds.). 1995. Neuroglia. Oxford University Press, New York,
оKuffler, S.W., and Nicholls, J. G. 1966. The physiology of neuroglial cells. Ergeb. Physiol. 57: 1-90.
оNewman, E., and Reichenbach, A. 1996. The Muller cell: A functional element of the retina. Trends Neurosci. 19: 307-312.
оPaulson, О. В., and Newman, E. A. 1987. Does the release of potassium from astrocyte endfeet regulate cerebral blood flow? Science 237: 896-898.
оPorter, J. T., and McCarthy, K. D. 1998. Astrocytic neurotransmitt r receptors in-situ and in-vivo. Prog. Neurobiol. 51: 439-455.
оRansom, B. R., and Orkand, R. K. 1996. Glialneuronal interaction in non-synaptic areas of the brain: Studies in the optic nerve. Trends Neurosci. 19: 352-358.
оRansom, B. R., and Sontheimer, H. 1992. The neurophysiology of glial cells. /. Clin. Neurophysiol.
glial cells. /. Clin. Neurophysiol.
9: 224-251.
о Saunders, N. R., Habgood, M. D., and Dziegielew-ska, К. М. 1999. Barrier mechanisms in the brain. I. Adult brain. Clin. Exp. Pharmacol. Physiol. 26: H-19.
Статьи
оAraque, Α., Li, N.. Doyle, R.T., and Haydon, P.O. 2000. SNARE protein-dependent glutmate release from astrocytes. /. Neurosci. 20: 666-673.
оKuffler, S.W., and Potter, D.D. 1964. Glia in the leech central nervous system: Physiological properties and neuron-glia relationship. /. Neurophysiol. 27: 290-320.
оNewman, Ε. Α., and Zahs, K. R. 1998. Modulation of neuronal activity by glial cells in the retina. /. Neurosci. 18: 4022-4028.
оPareek, S., Notterpek, L., Snipes, G. J., Naef, R., Sossin, W., Laliberte, J., lacampo, S., Suter, U, Shooter, E. M., and Murphy, R.A. 1997. Neurons promote the translocation of peripheral myelin protein 22 into myelin. /. Neurosci. 17: 7754-7762.