

Nikolls_-_Ot_neyrona_k_mozgu
.pdfГлава 20. Кодирование сигнала в латеральном коленчатом теле и первичной зрительной коре
Ганглиозные клетки сетчатки проецируют свои отростки в латеральное коленчатое тело, где они формируют ретинотопическую карту. У млекопитающих латеральное коленчатое тело состоит из 6 слоев, каждый из которых иннервируется либо одним, либо другим глазом и получает сигнал от различных подтипов ганглиозных клеток, образующих слои крупноклеточных (magnocellular), мелкоклеточных (parvocellular) и кониоклеточных*) (koniocellular) нейронов. Нейроны латерального коленчатого тела имеют рецептивные поля типа «центр-фон», подобно ганглиозным клеткам сетчатки.
Нейроны латерального коленчатого тела проецируются и формируют ретинотопическую карту в первичной зрительной коре V1, также называемой «зоной 17» или полосатой корой (striate cortex). Рецептивные поля кортикальных клеток, вместо уже привычной организации рецептивных полей по типу «центр-фон», состоят из линий, или краев, что является принципиально новым шагом в анализе зрительной информации. Шесть слоев V1 имеют особенности строения: афферентные волокна из коленчатого тела заканчиваются в основном в слое 4 (и некоторые в слое 6); клетки в слоях 2, 3 и 5 получают сигналы от кортикальных нейронов. Клетки слоев 5 и б проецируют отростки в подкорковые области, а клетки 2 и 3 слоя — в другие корковые зоны. Каждая вертикальная колонка клеток функционирует как модуль, получая исходный зрительный сигнал от определенного места в пространстве и посылая переработанную зрительную информацию во вторичные зрительные зоны. Колоночная организация зрительной коры очевидна, так как локализация рецептивных полей остается одинаковой на протяжении всей глубины коры, и зрительная информация от каждого глаза (правого или левого) всегда обрабатывается строго определенными колонками.
Было описано два класса нейронов в области V1, которые различаются по своим физиологическим свойствам. Рецептивные поля простых клеток удлинены и содержат сопряженные "on"- и "off''-зоны. Поэтому наиболее оптимальным стимулом для простой клетки является особым образом ориентированные пучки света или тени. Сложная клетка отвечает на определенным образом ориентированную полоску света; эта полоска может находиться в любой области рецептивного поля. Возникающее в результате распознавания изображения ингибирование простых или сложных клеток несет еще более детализированную информацию о свойствах сигнала, такую как наличие линии определенной длины или определенного угла в пределах данного рецептивного поля.
Рецептивные поля простой клетки образуются в результате конвергенции значительного количества афферентов из коленчатого тела. Примыкающие друг к другу центры нескольких рецептивных полей образуют одну корковую рецептивную зону. Поле сложной клетки зависит от сигналов простой клетки и других кортикальных клеток. Последовательное изменение организации рецептивных полей от сетчатки к латеральному коленчатому телу и затем к простым и сложным кортикальным клеткам говорит об иерархии в обработке информации, посредством чего ряд нейронных конструкций одного уровня интегрируется на следующем, где на основе исходной информации формируется еще более абстрактная концепция. На всех уровнях зрительного анализатора особое внимание уделяется контрастности и определению границ изображения, а не общей освещенности глаза. Таким образом, сложные клетки зрительной коры могут «видеть» линии, являющиеся границами прямоугольника, и их мало волнует абсолютная интенсивность света внутри этого прямоугольника.
*) Konio — греч. пыль, мельчайшие частички. — Прим перев.
444 |
Раздел III. Интегративные механизмы |
Серия четких и продолжающих друг друга исследований в области механизмов восприятия зрительной информации, начатая пионерскими работами Куффлера1) с сетчаткой, была продолжена на уровне зрительной коры Хьюбелем и Визелем. Хьюбель дал яркое описание ранних экспериментов на зрительной коре в лаборатории Стивена Куффлера в Университете Джона Хопкинса (США) в 50-х годах XX века2). С тех пор наше понимание физиологии и анатомии коры больших полушарий значительно развилось благодаря экспериментам Хьюбеля и Визеля, а также благодаря большому количеству работ, для которых их исследования были отправной точкой или источником вдохновения. Наша цель — дать краткое, повествовательное описание кодирования сигнала и архитектуры коры в аспекте восприятия, основанное на классических работах Хьюбеля и Визеля, а также на более поздних экспериментах, выполненных ими, их коллегами, а также многими другими. В этой главе мы лишь дадим схематический набросок функциональной архитектуры латерального коленчатого тела и зрительной коры, а также их роли в обеспечении первых шагов анализа зрительных сиен: определение линий и форм на основе поступающего из сетчатки сигнала в форме «центрфон».
При продвижении от сетчатки к латеральному коленчатому телу, а затем и к коре полушарий возникают вопросы, которые стоят вне пределов техники. В течение длительного время было общепризнанным, что для понимания функционирования любой части нервной системы необходимо знание о свойствах составляющих ее нейронов: каким образом они проводят сигналы и несут информацию, каким образом передают полученную информацию от одной клетки к другой посредством синапсов. Однако мониторинг активности только одной отдельной клетки вряд ли может быть результативным методом для изучения высших функций, где вовлечено большое количество нейронов. Аргумент, который здесь использовался и продолжает использоваться время от времени, следующий: мозг содержит около 1010 или более клеток. Даже самая простая задача или событие вовлекают сотни тысяч нервных клеток, расположенных в различных частях нервной системы. Каковы же шансы физиолога суметь проникнуть в суть механизма формирования сложного действия в головном мозге, если он может одновременно исследовать только одну или несколько нервных клеток, безнадежно малую долю от общего количества?
При более тщательном изучении логика подобных аргументов относительно основной сложности исследования, связанной с большим количеством клеток и сложными высшими функциями, уже не кажется такой безупречной. Как это часто происходит, появляется упрощающий принцип, открывающий новый и ясный взгляд на проблему. Ситуацию в зрительной коре упрощает то, что основные клеточные типы расположены отдельно друг от друга, в виде хорошо организованных и повторяющихся единиц. Эта повторяющаяся структура нервной ткани тесно переплетена с ретинотопической картой зрительной коры. Таким образом, соседние точки сетчатки проецируются на соседние точки поверхности зрительной коры. Это означает, что зрительная кора организована таким образом, чтобы для каждого мельчайшего сегмента зрительного поля находился набор нейронов для анализа информации и ее передачи. Кроме того, при помощи методов, которые позволяют выделить функционально связанные клеточные ансамбли, были выделены паттерны корковой организации более высокого уровня. В самом деле, архитектура коры определяет структурную основу корковой функции, поэтому новые анатомические подходы вдохновляют на новые аналитические исследования. Таким образом, прежде чем мы опишем функциональные связи зрительных нейронов, полезно вкратце резюмировать общую структуру центральных зрительных путей, начинающихся от ядер латерального коленчатого тела.
§ 1. Латеральное коленчатое тело
Волокна зрительного нерва начинаются от каждого глаза и заканчиваются на клетках правого и левого латерального коленчатого тела (ЛКТ) (рис. 20.1), имеющего четко различимую слоистую структуру («коленчатый» — geniculate — означает «изогнутый подобно колену»)3). В ЛКТ кошки можно увидеть три явных, хорошо различимых слоя клеток (А, А1, С), один из которых (А1) имеет сложное строение и подразделяется далее. У обезьян и других приматов, включая

Глава 20. Кодирование сигнала β латеральном коленчатом теле... |
445 |
|
|||||||
Рис. |
20.1. |
Латеральное |
|
|
|
|
|||
коленчатое тело (ЛКТ). (А) |
|
|
|
|
|||||
У кошки в ЛКТ имеется три |
|
|
|
|
|||||
слоя |
клеток: |
А, |
А, |
и |
С. |
|
|
|
|
(В) ЛКТ обезьяны имеет 6 |
|
|
|
|
|||||
основных |
|
|
слоев, |
|
|
|
|
||
включающих |
|
|
|
|
|
|
|
|
|
мелкоклеточные |
|
|
|
|
|
|
|
||
(рагvocellular), или Ρ (3, 4, |
|
|
|
|
|||||
5, 6), |
крупноклеточные |
|
|
|
|
||||
(magnocellular), или |
M |
(1, |
|
|
|
|
|||
2), |
разделенные |
|
|
|
|
||||
кониоклеточными |
|
|
|
|
|
|
|
||
(koniocellular) |
слоями (К). |
|
|
|
|
||||
У обоих животных каждый |
|
|
|
|
|||||
слой |
получает |
сигналы |
|
|
|
|
|||
только от одного глаза и |
|
Fig. 20.1. Lateral Geniculate Nucleus. (A) In cat LGN there are three layers of cells: A, |
|
||||||
содержит |
|
клетки, |
|
|
|||||
|
Al and С. (В) Monkey LGN has six major layers designated parvocellular, or Ρ (3, 4, 5, |
||||||||
имеющие |
|
|
|
|
|
||||
|
|
|
|
|
6) and magnocellular, or M (1, 2), separated by the koniocellular (K) layers. In both |
||||
специализированные |
|
|
|||||||
|
|
animals, each layer is supplied by only one eye and contains cells with specialized |
|||||||
физиологические |
|
|
|
|
|||||
|
|
|
|
response properties. (A from Szentâgothai, 1973; В after Hendry and Calkins, 1998.) |
|||||
свойства. |
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
||
человека, ЛКТ имеет шесть слоев клеток. Клетки в более глубоких слоях 1 и 2 больше по размерам, чем в слоях 3, 4, 5 и 6, из-за чего эти слои и называют соответственно
крупноклеточными (M, magnocellular) и мелкоклеточными (Р, parvocellular).
Классификация коррелирует также с большими (М) и маленькими (Р) ганглиозными клетками сетчатки, которые посылают свои отростки в ЛКТ. Между каждым M и Р слоями лежит зона очень маленьких клеток: интраламинарный, или кониоклеточный (К, koniocellular) слой. Клетки К слоя отличаются от M и Р клеток по своим функциональным и нейрохимическим4) свойствам, образуя третий канал информации в зрительную кору5).
Как у кошки, так и у обезьяны каждый слой ЛКТ получает сигналы либо от одного, либо от другого глаза. У обезьян слои 6, 4 и 1 получают информацию от контралатерального глаза, а слои 5, 3 и 2 — от ипсилатерального*). Разделение хода нервных окончаний от каждого глаза в различные слои было показано при помощи электрофизиологических и целого ряда анатомических методов6)--8). Особенно удивительным является тип ветвления отдельного волокна зрительного нерва при инъекции в него фермента пероксидазы хрена (рис. 20.2).
Образование терминалей ограничено слоями ЛКТ для этого глаза, без выхода за границы этих слоев. Из-за систематического и определенным образом проводимого разделения волокон зрительного нерва в районе хиазмы, все рецептивные поля клеток ЛКТ расположены в зрительном поле противоположной стороны (глава 19).
Рис. 20.2. Окончания волокон зрительного нерва в ЛКТ кошки. В один из аксонов от зоны с "on" центром контралатерального глаза была введена пероксидаза хрена. Веточки аксона заканчиваются на клетках слоев А и С, но не А1.
Fig. 20.2. Termination of an Optic Nerve Fiber in cat LGN. A single "on"-center axon from the contracterai eye was injected with horseradish peroxidase. Branches end in layers A and C, but not in A (After Bowling and Michael 1980.)
*) Гомолатерального, т е. с той же стороны. — Прим. перев.

446 |
Раздел III. Интегративные механизмы |
Рис. 20.3. Рецептивные поля клеток ШТ. Концентрические рецептивные поля клеток ЛКТ напоминают поля ганглиозных клеток в сетчатке, разделяясь на поля с "on"- и "off''-центром. Показаны ответы клетки с "on"-центром ЛКТ кошки. Полоской над сигналом показана продолжительность освещения. Центральные и периферические зоны нивелируют эффекты друг друга, поэтому диффузное освещение всего рецептивного поля дает только слабые ответы (нижняя запись), еще менее выраженные, чем в ганглиозных клетках сетчатки (сравните с рис. 19.21).
Fig. 20.3. Receptive Fields of Lateral Geniculate Nucleus Cells. The concentric receptive fields of cells in the LGN resemble those of ganglion cells in the retina, consisting of "on"-center and "off-center types. The responses illustrated are from an "on"-center cell in cat LGN. The bar above each record indicates illumination. The central and surround areas antagonize each other's effects, so diffuse illumination of the entire receptive field gives only weak responses (bottom record), less pronounced than in retinal ganglion cells (compare Figure 19.21). (After Hubel and Wiesel, 1961.)
Карты зрительных полей в латеральном коленчатом теле
Важной топографической особенностью является высокая упорядоченность в организации рецептивных полей в пределах каждого слоя ЛКТ. Соседние регионы сетчатки образуют связи с соседними клетками ЛКТ, так что рецептивные поля близрасположенных нейронов ЛКТ перекрываются на большой площади4· 5). Клетки центральной зоны сетчатки кошки (регион, где сетчатка кошки имеет маленькие по размеру рецептивные поля с малыми центрами), а также зрительной ямки обезьяны образуют связи с относительно большим количеством клеток в пределах каждого слоя ЛКТ. Подобное же распределение связей было обнаружено и у человека при помощи ЯМР9). Число клеток, связанных с периферическими регионами сетчатки, относительно мало. Такая избыточная представленность зрительной ямки отражает высокую плотность фоторецепторов в той зоне, которая необходима для зрения с максимальной остротой10). Хотя, наверное, число волокон зрительного нерва и число клеток ЛКТ примерно равны, тем не менее каждый нейрон ЛКТ получает конвергирующие сигналы от нескольких волокон зрительного нерва. Каждое волокно зрительного нерва в свою очередь образует дивергирующие синаптические связи с несколькими нейронами ЛКТ.
Однако каждый слой не только топографически упорядочен, но также и клетки разных слоев находятся в ретинотопическом отношении друг к другу. То есть, если продвигать электрод строго перпендикулярно к поверхности ЛКТ, то сначала будут регистрироваться активность клеток, получающих информацию от соответствующих зон одного, а затем и другого глаза, по мере того, как микроэлектрод пересекает один слой ЛКТ за другим. Расположение рецептивных полей находится в строго соответствующих позициях на обеих сетчатках, т. е. они представляют одну и ту же область зрительного поля4· 5). В клетках ЛКТ не происходит значительного смешивания информации от правого и левого глаза и взаимодействия между ними, лишь небольшое количество нейронов (которые имеют рецептивные поля в обоих глазах) возбуждаются исключительно бинокулярно.
Удивительно то, что ответы клеток ЛКТ не имеют разительных отличий от сигналов ганглиозных клеток (рис. 20.3). Нейроны ЛКТ также имеют концентрически организованные антагонизирующие рецептивные поля, либо с "off"-, либо с "on"-центром, но механизм контраста отрегулирован тоньше, за счет большего соответствия меж-
Глава 20. Кодирование сигнала в латеральном коленчатом теле... |
447 |
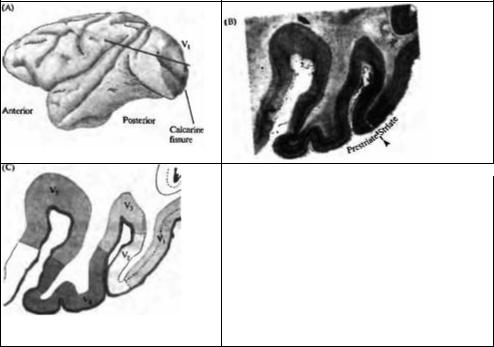
Рис. 20.4. Связь между первичной зрительной корой (V1) и V2 , V3 , V4 и V5 y обезьян. (А) Плоскость среза проходит через V, и V2 зоны. Четко видна граница между зонами V1 и V2. (В, С) Срез через затылочную кору. На В граница между V1 и V2 показана пунктирной линией в области, где исчезает полосатость. Границы между V2, V3, V4 и V5 зонами выявляются при помощи комбинации физиологических и морфологических исследований.
Fig. 20.4. Relation of Primary Visual Cortex (V1) to V2, V3, V4, and
V5 in the monkey. (A) The cortex and plane of section passing through V1 and V2. The boundary between V1 and V2 is unambiguous. (В, С) A section through the occipital cortex. In B, the boundary between V1 (striate) and V2 (prestriate) occurs at the dotted line, where the striped appearance is lost. The boundaries between V2, V3, V4, and V are revealed by a combination of physiological and anatomical studies. (After Zeki,
1990; micrograph kindly provided by S. Zeki and M. Rayan.)
ду тормозными и возбуждающими зонами. Таким образом, подобно ганглиозным клеткам сетчатки, для нейронов ЛКТ оптимальным стимулом является контраст, однако они реагируют еще слабее на общее освещение. Изучение рецептивных полей нейронов ЛКТ еще не завершено. Например, в ЛКТ найдены нейроны, вклад которых в работу ЛКТ не был установлен, а также пути, идущие от коры вниз к ЛКТ 11). Корковая обратная связь необходима для синхронизированной активности нейронов ЛКТ 12)--14).
Функциональные слои ЛКТ
Почему в ЛКТ на каждый глаз приходится больше одного слоя? Сейчас обнаружено, что нейроны в разных слоях имеют различные функциональные свойства. Например, клетки, находящиеся в четвертых дорзальных мелкоклеточных слоях ЛКТ обезьяны, подобно Ρ ганглиозным клеткам, способны отвечать на свет разных цветов, показывая хорошую цветовую дискриминацию15). И наоборот, слои 1 и 2 (крупноклеточные слои) содержат М- подобные клетки, которые дают быстрые («живые») ответы и нечувствительны к цвету, в то время как К слои получают сигналы от "blue-on" ганглиозных клеток сетчатки16) и могут играть особую роль в цветном зрении17). У кошек X и Y волокна (см. раздел «Классификация ганглиозных клеток» в главе 19) заканчиваются в различных подслоях А, С и А1, поэтому специфическая инактивация слоя А, но не С, резко снижает точность глазодвижений18). Клетки с "on"- и "off"-центром также подразделяются на различные слои в ЛКТ норки и хорька, и, в некоторой степени, у обезьян19)--21). Резюмируя вышесказанное, можно сказать, что ЛКТ является перевалочной станцией, в которой аксоны ганглиозных клеток сортируются таким образом, что соседние клетки получают сигналы от одинаковых регионов зрительных полей, и нейроны, перерабатывающие информацию, организованы в виде кластеров. Таким образом, в ЛКТ очевидной является анатомическая база для параллельной переработки (parallel processing) зрительной информации.

448 |
|
|
|
Раздел III. Интегративные механизмы |
Рис. |
20.5. |
Карта |
|
|
коркового |
|
зрительного |
|
|
поля. (А) Зрительное поле |
|
|||
разделяется |
|
на |
|
|
центральную |
|
зону |
|
|
(центральная ямка) и |
|
|||
восемь |
полуквадрантов. |
|
||
Левое зрительное |
поле |
|
||
проецируется |
на правую |
|
||
первичную |
зрительную |
|
||
кору. (В) |
У |
человека |
|
|
первичная |
зрительная |
|
||
кора («зона 17») почти |
|
|||
полностью |
лежит |
на |
|
|
медиальной |
поверхности |
непропорционально большую долю первичной зрительной коры. |
||
затылочной |
|
доли. |
||
Представительство |
|
Fig. 20.5. Visual Field Map of the Cortex. (A) The visual field is divided into a central |
||
центральной |
|
янки |
||
расположено |
сзади, а |
zone (the fovea), and eight hemiquadrants. The left visual field maps onto the right |
||
периферические |
зоны |
primary visual cortex. (B) In humans, the primary visual cortex (Area 17) lies almost |
||
проецируются |
спереди. |
entirely on the medial surface of the occipital lobe. The foveal region is found most |
||
Ямка занимает |
|
posteriorly, with the peripheral zones mapping anteriorly. The fovea claims a |
||
|
disproportionate share of the primary visual cortex. |
|||
|
|
|
|
|
§2. Цитоархитектоника зрительной коры
Зрительная информация поступает в кору и ЛКТ через оптическую радиацию. У обезьян оптическая радиация заканчивается на складчатой пластинке, толщиной около 2 мм (рис. 20.4). Этот регион мозга — известный как первичная зрительная кора, зрительная зона 1 или V1 — также называется полосатой корой, или «зоной 17». Более старая терминология базировалась на анатомических критериях, разработанных еще в начале XX века. V1 лежит сзади, в области затылочной доли, и может быть распознана при поперечном разрезе по своему особому внешнему виду. Пучки волокон в этой области формируют полоску, ясно видную невооруженным глазом (поэтому зона и называется «полосатой», рис. 20.4В). Соседние зоны вне зоны полосатости также связаны со зрением. Зона, непосредственно окружающая зону V, называется зоной V2 (или «зона 18») и получает сигналы из зоны V, (см. рис. 20.4С). Четкие границы так называемой экстрастриарноq зрительной коры (V2-V5) нельзя установить при помощи визуального исследования мозга, хотя для этого разработан ряд критериев22)--26). Например, в V2 полосатая исчерченность исчезает, большие клетки расположены поверхностно, и грубые, косо расположенные миелиновые волокна видны в более глубоких слоях. Как будет показано в главе 21, различные зрительные зоны производят анализ разного типа.
Каждая зона имеет собственное представление зрительного поля сетчатки, спроецированное строго определенным, ретинотопическим образом. Карты проекций были составлены еще в эпоху, когда не было возможно проводить анализ активности отдельных клеток. Поэтому для картирования использовалось освещение пучками света небольших участков сетчатки и регистрация активности коры при помощи большого электрода. Эти карты, а также их современные аналоги, составленные недавно при помощи методов визуализации головного мозга, таких как позитронно-эмиссионная томография и функциональный ядерно-магнитный резонанс, показали, что площадь коры, отведенная на представление центральной ямки, гораздо больше по размерам, чем площадь, отведенная на всю остальную сетчатку27)--30). Эти находки, в принципе, соответствовали ожиданиям, поскольку распознавание образов корой осуществляется в основном за счет переработки информации от плотно расположенных в зоне ямки фоторецепторов. Такое представление аналогично расширенному представлению руки и лица в области первичной соматосенсорной коры (глава 18). Ямка сетчатки проецируется в затылочный полюс коры больших полушарий. Карта периферии сетчатки распространяется в переднем направлении вдоль медиальной поверхности затылочной доли (рис. 20.5)31). Из-за перевернутой картины, образуемой на сетчатке при помощи хрусталика, верхнее зрительное поле проецируется на нижнюю область сетчатки и пе-

Глава 20. Кодирование сигнала в латеральном коленчатом теле... |
449 |
Рис. 20.6. Архитектоника зрительной коры. (А) Четко различима слоистость при срезе через стриарную кору макаки (окраска по Нисслю, выявляющая тела нейронов). Волокна, идущие от ЛКТ, заканчиваются в слоях 4А, 4В и 4С. (В) Рисунок пирамидных и звездчатых клеток в зрительной коре кошки. Отростки, окрашенные по методу Гольджи расходятся большей частью радиально в толщу коры и проходят лишь на небольшие расстояния в латеральном направлении. (С) Рисунок с фотографии пирамидной клетки и отростчатой звездчатой клетки в коре кошки, инъецированной пероксидазой хрена после регистрации ее активности. Обе клетки являются простыми.
Fig. 20.6. Architecture of Visual Cortex. (A) Distinct layering of cells in a section of striate cortex of the macaque monkey, stained to show cell bodies (Nissl stain). Fibers arriving from the LGN end in layers 4A, 4B, and 4C. (B) Drawing of pyramidal and stellate cells in cat visual cortex. The processes, stained with Golgi technique, for the most part run radially through the thickness of the cortex and extend for relatively short distances laterally. (C) Drawing from photographs of a pyramidal cell and a spiny stellate cell in cat cortex that had been injected with horseradish peroxidase after their activity had been recorded.
Both were simple cells. (A from Hubel and Wiesel 1972; В after Ramon у Cajal 1955; С from Gilbert and Wiesel 1979.)
редается в область V1, расположенную ниже шпорной борозды; нижнее зрительное поле проецируется над шпорной бороздой.
На срезах коры нейроны могут быть классифицированы по их форме. Две основные группы нейронов образуют звездчатые и пирамидные клетки. Примеры этих клеток показаны на рис. 20.6В. Основные различия между ними заключаются в длине аксонов и в форме тел клеток. Аксоны пирамидных клеток длиннее, спускаются в белое вещество, покидая кору; отростки же звездчатых клеток заканчиваются в ближайших зонах. Эти две группы клеток могут иметь и другие различия, такие как наличие или отсутствие шипиков на дендритах, которые обеспечивают их функциональные свойства32). Есть и другие, причудливо названные нейроны (двухбукетные клетки, клетки-люстры, корзинчатые клетки, клетки-полумесяцы), а также клетки нейроглии. Их характерной особенностью является то, что отростки этих клеток направляются в основном в радиальном направлении: вверх и вниз через толщу коры (под соответствующим утлом к поверхности). И наоборот, многие (но не все) их латеральные отростки короткие. Соединения между первичной зрительной корой и корой высшего порядка осуществляется при помощи аксонов, которые проходят в виде пучков через белое вещество, находящееся под клеточными слоями.
450 |
Раздел III. Интегративные механизмы |
|
|
|
|
|
|
|
|
|
|
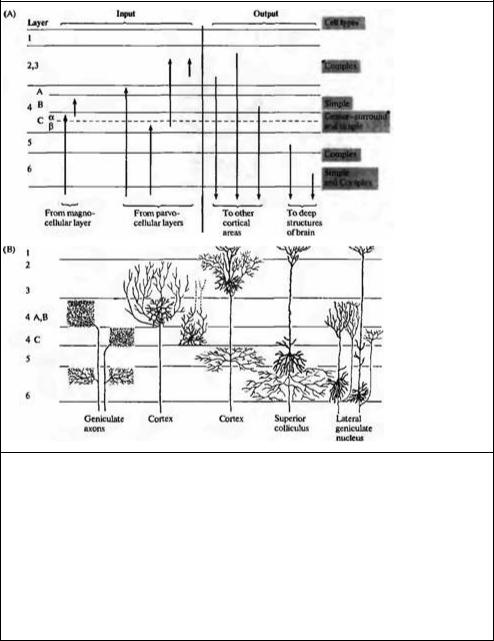
Рис. 20.7. Связи зрительной коры. (А) Слои клеток с различными входящими и исходящими отростками. Отметим, что исходные отростки из ЛКТ в основном прерываются в 4 слое. Отростки из ЛКТ, идущие от крупноклеточных слоев, преимущественно прерываются в 4Сα и 4В слоях, в то время как отростки от мелкоклеточных прерываются в 4А и 4Сβ. Простые клетки расположены в основном в слоях 4 и 6, комплексные клетки — в слоях 2, 3, 5 и 6. Клетки слоев 2, 3 и 4В посылают аксоны в другие корковые зоны; клетки в слоях 5 и 6 посылают аксоны к верхнему холмику и ЛКТ. (В) Типичная ветвистость аксонов ЛКТ и кортикальных нейронов кошки. Кроме подобных вертикальных связей, многие клетки имеют длинные горизонтальные связи, идущие в пределах одного слоя к удаленным регионам коры.
Fig. 20.7. Connections of Visual Cortex. (A) The layers with their various inputs, outputs, and types of cells. Note that inputs from the LGN end mainly in layer 4. Those arising from magnocellular layers end principally in 4Cα and 4B, whereas those from parvocellular layers end in 4A and 4Cβ. Simple cells are found mainly in layers 4 and 6, complex cells in layers 2, 3, 5, and 6. Cells in layers 2,3, and 4B send axons to other cortical areas; cells in layers 5 and 6 send axons to the superior colliculus and the lateral geniculate nucleus. (B) Principal arborizations of gemculate axons and cortical neurons in the cat. In addition to these vertical connections, many cells have long horizontal connections running within a layer to distant regions of the cortex. (A after Hubet 1988; В after Gilbert and WieseL 1981.)
Входящие, исходящие пути и послойная организация коры
Основной особенностью коры млекопитающих является то, что клетки здесь расположены в виде 6 слоев в пределах серого вещества (рис. 20.6А). Слои сильно различаются по внешнему виду, в зависимости от плотности расположения клеток, а также толщины каждой из зон коры. Входящие пути показаны на рис. 20.7A с левой стороны. Исходя из ЛКТ, волокна в основном заканчиваются в слое 4,
Глава 20. Кодирование сигнала в латеральном коленчатом теле... |
451 |
с небольшим количеством связей, образуемых также в слое 6. Поверхностные слои получают сигналы из области подушки таламуса (pulvinar zone) или других зон таламуса. Большое количество клеток коры, особенно в области слоя 2, а также в верхних частях слоев 3 и 5 получают сигналы от нейронов, также расположенных в пределах коры. Основная масса волокон, идущих от ЛКТ в слой 4, затем разделяется между различными подслоями, детали строения и функциональное значение которых будет нами рассмотрено в главе 21.
Исходящие из слоев 6, 5, 4, 3 и 2 волокна показаны справа на рис. 20.7А11, 33). Клетки, посылаюшие эфферентные сигналы из коры, могут также управлять внутри корковыми соединениями между разными слоями. Например, аксоны клетки из слоя 6, кроме ЛКТ, могут также направляться в один из других кортикальных слоев, в зависимости от типа ответа этой клетки34). На основании подобного строения зрительных путей можно представить следующий путь зрительного сигнала: информация с сетчатки передается на клетки коры (в основном, в слой 4) аксонами клеток ЛКТ; информация передается из слоя в слой, от нейрона к нейрону по всей толщине коры; переработанная информация пересылается в другие зоны коры при помощи волокон, направляющихся вглубь белого вещества и возвращающихся обратно в область коры. Таким образом, радиальная или вертикальная организация коры дает нам основания полагать, что колонки нейронов работают как отдельные вычислительные единицы, обрабатывая различные детали зрительных сцен и пересылая полученную информацию далее в другие регионы коры.
Разделение входящих волокон от ЛКТ в слое 4
Афферентные волокна ЛКТ заканчиваются в слое 4 первичной зрительной коры, который имеет сложную организацию и может быть исследован как физиологически, так и анатомически. Первой особенностью, которую мы хотим продемонстрировать, является разделение входящих волокон, идущих от разных глаз. У взрослых кошек и обезьян клетки в пределах одного слоя ЛКТ, получая сигналы от одного глаза, посылают отростки к строго определенным скоплениям клеток коры в слое 4С, отвечающим именно за этот глаз. Скопления клеток сгруппированы в виде чередующихся полосок или пучков кортикальных клеток, получающих информацию исключительно от правого или левого глаза. В более поверхностно и глубже расположенных слоях нейроны управляются обоими глазами, хотя обычно с преобладанием одного из них. Хьюбель и Визель провели оригинальную демонстрацию разделения информации от разных глаз и преобладания одного из них в первичной зрительной коре при помощи электрофизиологических методов. Они использовали термин «глазодоминантные колонки» (ocular dominance columns) при описании своих наблюдений, придерживаясь концепции кортикальных колонок, разработанной Маунткаслом для соматосенсорной коры (глава 18)35). Серия экспериментальных методик была разработана для демонстрации чередующихся групп клеток в слое 4, получающих информацию от правого или левого глаза. Вначале было предложено нанести небольшое повреждение в пределах только одного слоя ЛКТ (напомним, что каждый слой получает информацию только от одного глаза). Если это сделать, то дегенерирующие терминали появляются в слое 4, образуя определенный паттерн чередующихся пятен, которые соответствуют зонам, управляемым глазом, посылающим информацию в поврежденную область ЛКТ. Позднее потрясающая демонстрация существования особого паттерна глазного доминирования была выполнена, используя транспорт радиоактивных аминокислот из одного глаза. Эксперимент состоит в том, что в глаз вводится аминокислота (пролин или лецитин), содержащая атомы радиоактивного трития. Инъекция проводится в стекловидное тело глаза, из которого аминокислота захватывается телами нервных клеток сетчатки и включается в состав белка. Со временем помеченный таким образом белок транспортируется в ганглиозные клетки и по волокнам зрительного нерва в их терминали в пределах ЛКТ. Замечательной особенностью является то, что эта радиоактивная метка также передается от нейрона к нейрону через химические синапсы36)--38). В конечном итоге метка попадает в окончания волокон ЛКТ в пределах зрительной коры.
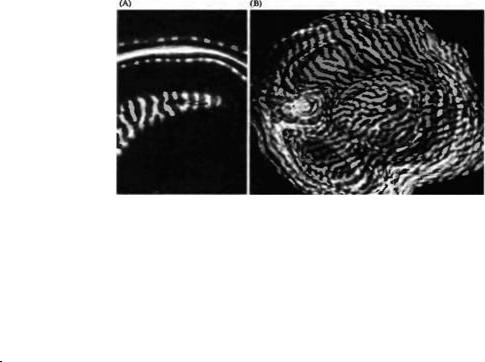
На рис. 20.8 показано расположение в пределах слоя 4 радиоактивных терминалей, образованных аксонами клеток ЛКТ, связанных с глазом, в который вводилась метка (радиоактивность вызывает отложение зерен серебра на фотографической пленке, которую
452 |
|
|
|
|
Раздел III. Интегротивные механизмы |
|
Рис. |
|
|
20.8. |
|
|
|
Глазодоминантные |
|
|
|
|||
колонки |
|
в |
|
коре |
|
|
обезьяны, |
полученные |
|
|
|||
при помощи введения |
|
|
||||
радиоактивного |
|
про |
|
|
||
лина в один глаз. |
|
|
||||
Ауторадиограммы, |
|
|
|
|||
снятые |
|
|
|
при |
|
|
темнопольном |
|
|
|
|
||
освещении, |
где |
белым |
|
|
||
показаны зерна серебра. |
|
|
||||
(А) Сверху рисунка срез |
|
|
||||
проходит через |
слой 4 |
|
|
|||
зрительной |
коры |
под |
|
|
||
углом |
к |
поверхности, |
|
|
||
образуя |
|
|
|
|
|
|
перпендикулярный |
срез |
|
|
|||
колонок. В центре слой 4 |
лишь часть слоя 4, что обусловлено кривизной коры.) |
Как в А, так и в В колонки |
||||
был |
|
|
срезан |
|||
горизонтально, |
|
|
зрительного доминирования выглядят как полоски равной ширины, получающие |
|||
показывая, |
что |
колонка |
информацию либо от одного, либо другого глаза. |
|
||
состоит |
из |
удлиненных |
Fig. 20.8. Ocular Dominance Columns in monkey cortex demonstrated by injection of |
|||
пластинок. |
|
|
(В) |
|||
Реконструкция |
|
из |
radioactive praline into one eye. Autoradiographs photographed with dark-field |
|||
множества |
|
|
|
illumination in which the silver grains appear white. (A) |
At the top of the picture the |
|
|
|
|
section passes through layer 4 of the visual cortex at |
right angles to the surface, |
||
горизонтальных |
срезов |
|||||
слоя |
4С |
у |
другой |
displaying columns cut perpendicularly. In the center, layer 4 has been cut horizontally, |
||
обезьяны, |
у |
которой |
showing that the columns consist of longer slabs. (B) Reconstruction made from |
|||
инъекция проводилась в |
numerous horizontal sections of layer 4C in another monkey in which the ipsilateral eye |
|||||
илсилатеральный |
глаз. |
had been injected. (No single horizontal section can encompass more than a part of layer |
||||
(Любой |
горизонтальный |
4 because of the curvature of the cortex.) In both A and B, the ocular dominance columns |
||||
разрез может выявить |
appear as stripes of equal width supplied by one eye or the other. (Autoradiographs |
|||||
kindly provided by S. LeVay.) |
|
|||||
|
|
|
|
|
|
|
располагают непосредственно над зрительной корой, поэтому такие участки выглядят как белые пятна на темном фоне фотографии). Пятна от меток перемежаются с зонами без меток, которые получают информацию от контралатерального глаза, куда не вводилась метка. Расстояние от центра до центра между пятнами, которые соответствуют глазодоминантным колонкам, составляет приблизительно 1 мм.
На клеточном уровне сходная структура была выявлена в слое 4 при помощи введения пероксидазы хрена в отдельные направляющиеся в кору аксоны нейронов ЛКТ39)--40). Аксон, показанный на рис. 20.9, идет от нейрона ЛКТ с "off"-центром, отвечающим короткими сигналами на тени и движущиеся пятна. Аксон заканчивается в двух различных группах отростков в слое 4. Группы меченых отростков отделены пустой немеченной зоной, соответствующей по своим размерам территории, отвечающей за другой глаз. Подобного рода морфологические исследования расширяют границы и позволяют более глубоко понять оригинальное описание колонок глазного доминирования, составленное Хьюбелем и Визелем в 1962 году.
§ 3. Стратегии изучения коры
Проблемой, стоящей перед Хьюбелем и Визелем в 1958 году было то, каким образом сигналы, определяющие маленькие, яркие, темные или цветовые пятна на сетчатке могут быть преобразованы в сигналы, которые несут информацию о форме, размере, цвете, движении и глубине расположения объектов. Методики, считающиеся сейчас рутинными, такие как оптическая регистрация, введение пероксидазы хрена или сканирование мозга, в ту пору даже не могли быть представлены. В начале работы перед Хьюбелем и Визелем стояли совершенно неизученные вопросы, которые исходили из их предположения, что зрительные центры в коре осуществляют переработку информации подобно тому, как это происходит в сетчатке, только на более высоком уровне.
Одной из ключевых стратегий в их анализе было использование стимулов, которые имитировали сигналы, возникающие в нормальных условиях. Например, края, контуры, а также простые паттерны, представленные глазу, позволяли выявлять особенности организации, которые никогда бы не были об-