

Nikolls_-_Ot_neyrona_k_mozgu
.pdfГлава 22. Клеточные механизмы двигательного контроля |
493 |
|
|||||
Рис. 22.6. Организация синаптических |
|
|
|
||||
контактов для рефлексов спинного мозга. |
|
|
|
||||
Спинной мозг показан в поперечном |
|
|
|
||||
сечении, с указанием интернейронов. (А) |
|
|
|
||||
При миотатическом рефлексе растяжение |
|
|
|
||||
мышечных веретен генерирует импульсы, |
|
|
|
||||
которые проходят no la афферентным |
|
|
|
||||
волокнам в спинной мозг и вызывают |
|
|
|
||||
моносинаптическое |
возбуждение |
в |
α |
|
|
|
|
мотонейронах той же мышцы. Импульсы |
|
|
|
||||
возбуждают также интернейроны, которые, в |
|
|
|
||||
свою очередь, ингибируют мотонейроны, |
|
|
|
||||
снабжающие |
мышцы-антагонисты. |
(В) |
|
|
|
||
Активация сухожильных органов |
Гольджи |
|
|
|
|||
вызывает импульсацию в Ib афферентах, |
|
|
|
||||
которые, |
через |
интернейроны, |
|
|
|
||
обеспечивают |
торможение мотонейронов |
|
|
|
|||
этой мышцы и одновременно возбуждают |
|
|
|
||||
мотонейроны мышц-антагонистов. |
|
|
|
|
|
||
Fig. 22.6. Organization of Synaptic Connections |
|
|
|
||||
for reflex actions in the spinal cord. The spinal |
|
|
|
||||
cord is shown in transverse section, with |
|
|
|
||||
inhibitory interneurons in black. (A) In the |
|
|
|
||||
myotatic reflex, stretch of the muscle spindle |
(B) Activation of Golgi tendon organs produces |
impulses in group Ib |
|||||
generates impulses that travel along group la |
|||||||
afferent fibers to the spinal cord and produce |
afferent fibers that through interneuronal |
connections, provide |
|||||
monosynaptic excitation of Q-motoneurons to |
inhibition to motoneurons supplying the same |
muscle and excitation |
|||||
that same muscle. Impulses also excite |
to antagonist motoneurons. (This is sometimes |
called the inverse |
|||||
interneurons that in turn, inhibit motoneurons |
myotatic reflex.) |
|
|
||||
supplying the antagonist muscles. |
|
|
|
|
|
||
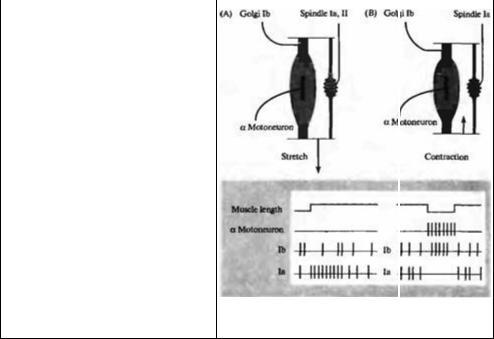
вать потенциал действия. Принцип размера справедлив для всех синаптических контактов, поскольку они равномерно распределены среди мотонейронов12)--14).
§ 2. Спинальные рефлексы Реципрокная иннервация
Движение конечностей обусловлено координированным сокращением работающих вместе мышц, называемых агонистами. В это же время мышцы противоположного функционального назначения, называемые антагонисты, находятся в состоянии расслабления. Мышцы-удлинители конечностей разгибают или удлиняют суставы, что помогает преодолеть силы притяжения, мышцы-сгибатели сгибают суставы и притягивают конечности к телу. При активации миотатического рефлекса растяжением мышцы, например при ударе по коленной чашечке, первичные сенсорные окончания в мышечных веретенах мышц--удлинителей (четырехглавой мышце бедра) подвергаются деформации и инициируют поток импульсов в Ia афферентах, направляющихся в спинной мозг. Эти импульсы продуцируют моносинаптическое возбуждение в α-мотонейронах, посылающих сигналы обратно к растянутой мышце, что, в конечном счете, приводит к ее рефлекторному сокращению. Сокращение мышцы-удлинителя сопровождается одновременным торможением α-мотонейрона, который иннервирует антагонистическую мышцу-сгибатель. Это происходит потому, что Ia афференты активируют также интернейроны, которые, в свою очередь, ингибируют α-мотонейроны антагонистов (рис. 22.6А). Принцип, когда активация одной группы мышц сопровождается торможением антагонистов, был впервые описан Шерринггоном, который назвал его реципрокной иннервацией.
Для простоты, на рис. 22.6А не показаны некоторые проводящие пути. Например, импульсы от небольшой группы II афферентов усиливают рефлексы как напрямую через моносинаптические контакты, так и посредством подключения интернейронов15). Эти внутриспинальные переключения были описаны в деталях16)—18). Ингибиторные интернейроны активируются также сухожильными органами Гольджи (СОГ). чувствительные окончания которых инкапсулированы на месте соединений мышцы и сухожилия (рис. 22.6В). Их афферентные волокна обо-
494 |
|
|
|
|
|
|
Раздел III. Интегративные механизмы |
Рис. |
22.7. |
|
Отслеживание |
мышечного |
|||
сокращения. (А) Во время внешнего |
|||||||
растяжения |
экстра |
фуэальных |
|
мышечных |
|||
волокон la афференты реагируют на него |
|||||||
благодаря |
|
своему |
параллельному |
||||
расположению. В отличие от этого, Ib |
|||||||
афференты |
сухожильных |
органов |
Гольджи |
||||
отвечают очень слабо либо не отвечают |
|||||||
вообще, благодаря их последовательному |
|||||||
расположению |
и |
присущей |
|
мышцам |
|||
эластичности. (В) Во время активации α |
|||||||
мотонейронов |
|
экс |
трафузальные |
волокна |
|||
сокращаются, что приводит к возбуждению Ib |
|||||||
афферентов сухожильных органов Гольджи, |
|||||||
тогда как мышечные веретена не реагируют, |
|||||||
поскольку |
|
растяжение |
интрафузальных |
||||
волокон нейтрализовано укорочением мышцы. |
|||||||
Fig. 22.7. Sensing Muscle Contraction. (A) During an imposed stretch of the extrafusal muscle, the spindle's la afferent senses the imposed stretch directly because of its "parallel" arrangement. However, the Golgi tendon organ Ib afférents respond only slightly or not at alt because of their "serial" position and the inherent elasticity of muscle. (B) During activation of the a- motoneurons, the extrafusal muscle contracts and the Golgi tendon organ's Ib afférents are excited, but the spindle afférents quiet as the stretch of the intrafusal fibers is relieved by shortening of the muscle.
значаются как Ib, для того чтобы отличать их от афферентных волокон мышечных веретен. СОГ располагаются последовательно относительно сокращающихся мышечных волокон. Поэтому СОГ можно активировать пассивным растяжением мышцы, но адекватным раздражителем является мышечное сокращение. Достаточно сокращения всего одного или двух мышечных волокон, развивающих усилие меньше 100 мг, для того, чтобы вызвать ощутимый разряд в Ib афферентах19· 20). Эти волокна от СОГ активируют интернейроны, которые, в свою очередь, тормозят мотонейроны, иннервирующие мышцу, в которой были активированы Ib афференты21) (рис. 22.6В). Информация, получаемая от СОГ, позволяет выработать адекватную двигательную команду, которая передается в высшие моторные центры22).
Сенсорная информация от мышечных рецепторов
СОГ и мышечные веретена производят противоположный эффект на мотонейроны той мышцы, в которой они располагаются. Активность мышечных веретен возбуждает мотонейрон, тогда как СОГ через интернейроны оказывают тормозящее действие на мотонейроны. Различное расположение этих двух рецепторов обеспечивает не только быстрые рефлекторные реакции, но и снабжает мозг важной информацией о положении конечностей и силе мышечного сокращения (рис. 22.7). Как уже отмечалось ранее, за счет последовательного расположения СОГ относительно мышечного волокна они активируются при сокращении и слабо реагируют на внешние стимулы. Наоборот, мышечные веретена располагаются параллельно экстрафузальным мышечным волокнам и их активность снижается при мышечном сокращении. Мышечные веретена высоко чувствительны и к пассивному растяжению. В результате мышечные веретена служат как датчики длины, тогда как СОГ контролируют силу мышечных сокращений. Поскольку информация от той и другой афферентной систем направляется к высшим центрам, анализ этой сигнализации позволяет осуществлять тонкую регулировку мышечных сокращений.
Глава 22. Клеточные механизмы двигательного контроля |
495 |
|||||||||
Рис. |
22.8. |
Эфферентная |
|
|
||||||
регуляция |
|
|
мышечных |
|
|
|||||
веретен. |
|
|
|
|
(А) |
|
|
|||
Интрафузальные |
мышечные |
|
|
|||||||
волокна |
|
|
|
получают |
|
|
||||
иннервацию |
|
|
|
|
|
|
|
|||
γ?эфферентными волокнами, |
|
|
||||||||
которые, |
|
|
|
|
будучи |
|
|
|||
активированными, |
вызывают |
|
|
|||||||
сокращение |
интрафузальных |
|
|
|||||||
мышечных |
волокон. |
Это |
|
|
||||||
сокращение |
|
растягивает |
|
|
||||||
чувствительные |
|
окончания |
|
|
||||||
веретен, |
вызывая |
в |
них |
|
|
|||||
импульсацию |
|
|
|
(В) |
|
|
||||
Внеклеточное |
|
отведение |
от |
|
|
|||||
мышечного |
веретена |
в |
|
|
||||||
инспираторной |
мышце |
во |
|
|
||||||
время |
дыхательного |
цикла. |
|
|
||||||
(С) |
После |
избирательного |
|
|
||||||
блокирования |
фузимоторных |
|
|
|||||||
волокон |
|
|
|
прокаином |
|
|
||||
мышечные веретена |
ведут |
|
|
|||||||
себя |
пассивно, |
увеличивая |
|
|
||||||
разряды |
во |
время |
выдоха |
|
|
|||||
(когда мышцы растянуты) и |
|
|
||||||||
прекращая |
активность |
во |
|
|
||||||
время вдоха, когда мышцы |
|
|
||||||||
укорочены. |
|
|
|
|
|
|
|
|||
Fig. 22.8. Efferent Regulation of |
|
|
||||||||
Muscle |
Spindles. (A) Intrafusal |
|
|
|||||||
muscle |
fibers |
of |
mammalian |
|
|
|||||
spindles receive |
innervation |
by |
|
|
||||||
γ efferent fibers that when |
|
|
||||||||
activated, |
cause |
the intrafusal |
|
|
||||||
fibers |
to |
contract. |
This |
|
|
|||||
contraction distorts the sensitive |
|
|
||||||||
endings of the spindle afférents, |
(B) Extracellular recording from a muscle spindle in an inspiratory muscle during the |
|||||||||
causing them to fire. The mass |
respiratory cycle (lower trace). Normally the sensory discharge from inspiratory |
|||||||||
of |
extrafusal |
muscle fibers |
is |
muscle spindles is highest during inspiration, even though the muscles are |
||||||
drawn |
separately; |
when |
it |
shortening. This is due to simultaneous activation of 7 efferents to the spindle. (C) |
||||||
contracts, |
stretch |
on |
the |
After the fusimotor fibers are blocked selectively by procaine, the spindles behave |
||||||
intrafusal fibers is reduced. |
|
passively, with increased discharges during expiration, when the muscles are |
||||||||
|
|
|
|
|
|
|
|
|
stretched, and cessation of activity on inspiration, when the muscles are shortened. |
|
|
|
|
|
|
|
|
|
|
(After Critchlow and von Euler, 1963.) |
|
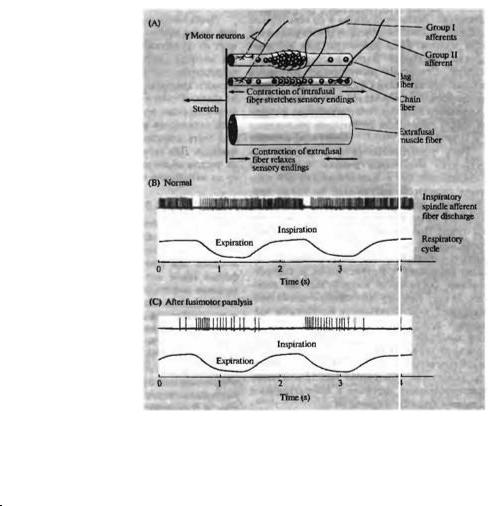
Эфферентный контроль мышечных веретен
Ответ от мышечных веретен осложнен тем, что веретена имеют собственные сократительные элементы, так называемые интрафузалъные мышечные волокна (глава 17). Они по ряду признаков отличаются от экстрафузальных волокон, которые образуют рабочую мышцу23). Так, они сокращаются в ответ на возбуждение, приходящее от небольших (диаметр 2-8 мкм) γ-мотонейронов по фузимоторным волокнам (рис. 22.8А). Фузимоторные волокна впервые были описаны Экклсом и Шеррингтоном24) и изучены в деталях Лекселлом25).
Роль фузимоторных волокон была окончательно установлена в серии технически сложных экспериментов, проведенных Куффлером с соавторами26). Суть этих экспериментов заключалась в регистрации активности отдельных афферентных Ia волокон дорзальных корешков, приходящих от мышечного веретена в ответ на стимуляцию фузимоторного волокна вентральных корешков, иннервирующего это же веретено. Стимуляция фузимоторного волокна приводила к усилению сенсорной активности, но не меняла мышечное на-
496 |
Раздел III. Интегротивные механизмы |
пряжение. Залпы импульсов в фузимоторном нейроне усиливали сенсорную импульсацию, если мышца была растянута, или инициировали сенсорный поток в сократившейся мышце. Сенсорная импульсация возникала потому, что активация фузимоторного нейрона приводила к сокращению интрафузального мышечного волокна, что растягивало и активировало окончания рецепторов растяжения I и II типа.
Роль γ-мотонейронов в регуляции импульсации мышечных веретен заключается в следующем: без γ-мотонейронального контроля рецепторы растяжения во время мышечного укорочения были бы неактивными и неспособными отслеживать изменения длины мышц. Поэтому во время сокращения экстрафузальных мышечных волокон, активируемых α-мотонейронами, параллельно происходит активация γ-мотонейронов и, соответственно, сокращение интрафузальных волокон. За счет этого фузимоторная система поддерживает чувствительность рецепторов мышечных веретен при любом объеме движений конечностей. Таким образом, фузимоторная иннервация мышечных веретен может рассматриваться как своеобразный «регулятор усиления», постоянно подстраивающий чувствительность датчиков к изменяющимся условиям функционирования.
Экспериментальные свидетельства совместной активации α- и γ-мотонейронов были получены в опытах27)--29), в которых отводились разряды мышечных веретен в инспираторных мышцах во время дыхательных движений. Рис. 22.8В показывает, что афферентный разряд в рецепторах растяжения инспираторных мышц, действительно, наиболее выражен во время вдоха, когда эти мышцы максимально укорочены. Это может получить объяснение, если допустить, что α- и γ-мотонейроны совместно активированы, так что сокращение интрафузальных волокон компенсирует и даже превышает укорочение экстрафузальных. Если же γ-мотонейроны блокированы местным анестетиком (рис. 22.8С), афферентные волокна возбуждаются только во время выдоха, который приводит к растяжению инспираторных мышц. Другое доказательство ко-активации α- и γ-мотонейронов было получено на модели движения пальца, когда разряды в мышечных афферентах отводились во время произвольных изометрических сокращений, не сопровождаемых движениями в суставах30).
Достаточно ясно, что наличие собственной иннервации мышечных веретен осложняет их функцию как простых датчиков длины мышцы (рис. 22.8). Разряды мышечных афферентов растянутых мышц без фузимоторного контроля могут быть такими же, как от укороченных мышц с активным участием фузимоторных волокон. Поэтому электрическая активность веретен сама по себе не может быть абсолютным индикатором длины мышцы. Для вычисления этой информации «в чистом виде» необходимо вносить поправку на активность фузимоторных влияний.
Сгибательные рефлексы
Активация болевых рецепторов кожи может приводить к сложным движениям, вовлекающим не только несколько суставов, а, порой, и более чем одну конечность. Простейшая форма такой активности называется сгибательным рефлексом, поскольку она включает активацию мышц-сгибателей и торможение разгибателей, что в результате приводит к удалению данной конечности от источника раздражения. Точный характер ответа зависит от локализации раздражения и типа вовлеченных болевых и тактильных рецепторов31). Ответная реакция приводит к активации целой нейрональной сети, включающей возбуждающие и тормозные интернейроны (рис. 22.9). Результирующее направление движения зависит от взаимодействия между степенью активации мотонейронов сгибателей и торможения мотонейронов разгибателей. Одновременно с этим активируются разгибатели с контралатеральной стороны, что балансируется ингибированием сгибателей. Такой перекрестно-разгибательный рефлекс является адекватной реакцией, когда мы, например, наступаем на острый гвоздь. Он демонстрирует высокую комплексность координации движений на уровне спинного мозга.
§3. Генерация координированных движений
До настоящего момента мы говорили об относительно простых двигательных рефлексах, базирующихся на прямом возбуждении моторных единиц сигналом, приходящим от рецепторов растяжения в самой мышце, причем этот же сигнал, через интернейрон, инги-

Глава 22. Клеточные механизмы двигательного контроля |
497 |
Рис. 22.9. Сгибательный рефлекс, относящийся к защитным, вызван в данном случае наступанием на гвоздь. Возбуждение болевых Αδ−волокон приводит к поднятию бедра и сгибанию колена благодаря полисинаптическому возбуждению мотонейронов сгибателей и торможению мышц-удлинителей конечностей.
Fig. 22.9. The Flexor Reflex is a timb-withdrawal reflex, produced in this example by stepping on a tack. Excitation of Ai pain fibers results in elevation of the thigh (synaptic connections not shown) and flexing of the knee joint by polysynaptic excitation of flexor motoneurons and inhibition of extensors (black interneurons are inhibitory). Also not shown are contracterai connections that subserve extension of the opposite leg for support.
бирует активность мышц-антагонистов. Интересно выяснить, базируется ли на тех же принципах реципрокной иннервации переменная активация сгибателей и разгибателей при повторяющихся, ритмических движениях, таких как ходьба или дыхание? Скорее всего, это действительно так, поскольку такие движения сопровождаются реципрокной активацией и торможением мышц-антагонистов. Однако есть одно важное отличие, заключающееся в том, что если даже подобные (а может, и те же самые) интернейроны используются для простого рефлекса и ритмической двигательной активности, сенсорная обратная связь не является обязательной для движения. Ритмические паттерны двигательной активности могут регистрироваться даже в изолированной нервной системе как позвоночных, так и беспозвоночных32).
Генераторы центрального ритма
Если ритмические разряды двигательной активности не связаны с сенсорным притоком, какие же клеточные механизмы могут генерировать командные сигналы на выходе? Предполагается, что это происходит благодаря двум механизмам: (1) ритмическим изменениям возбудимости одиночных нейронов, называемых водителями ритма (пейсмекерные клетки); (2) синаптическому взаимодействию нервных клеток в нейрональной сети. Вполне возможно, что эти механизмы работают вместе, производя разные паттерны ритмической активности в нервной системе.
Ритмическая активность в пейсмекерных нейронах возникает благодаря осцилляциям мембранного потенциала. Такие осцилляции могут запускаться различными ионными механизмами, но наиболее часто они базируются на обратной связи между входом кальция в клетку и открытием кальций-активируемых калиевых ионных каналов33). Калиевые ионные каналы важны для регуляции временного хода и паттерна ритмической активности в нейронах. Например, особый вид калиевых каналов был найден в генерирующих ритмическую активность нейронах омара34). Пейсмекерные нейроны были найдены как у позвоночных, так и у беспозвоночных, включая такие нейроны, которые отвечают за циркадные ритмы в сетчатке улитки, следующие с периодом в 24 часа35) (см главу 15). Эксперименты, проведенные на изолированном спинном мозге, показали наличие интернейронов-генераторов ритма, имеющие характеристики, сходные с паттерном локомоции36)--39).
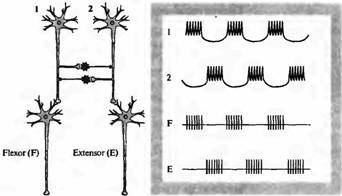
Достаточно ясно, что нейроны-водители ритма могут передать возбуждение непосредственно другим нейронам, включенным в сеть, например мотонейронам. Для обеспечения переключающегося возбуждения сгибателей и разгибателей требуется уже более сложная нейрональная сеть. Это может быть достигнуто по механизму, показанному на рис. 22.10, когда два идентичных пейсмекера (1 и 2) возбуждают, соответственно, мотонейрон сгибателя и разгибателя. Они же имеют тормозное влияние друг на друга, реализуемое через интернейроны. Благодаря взаимному торможению ни один из агонистов не может быть активным тогда, когда активен его антагонист. Таким образом, тормозные взаимодействия обеспечивают реципрокность функционирования групп нейронов. Генерация цен-
498 |
|
|
|
|
|
|
|
Раздел Ш. Интегративные механизмы |
Рис. |
22.10. |
|
Гипотетический |
|
||||
генератор |
ритма, |
состоящий |
|
|||||
из двух |
пейсмекерных клеток |
|
||||||
(1 и |
2), |
которые |
возбуждают |
|
||||
сгибатели |
и |
разгибатели, |
|
|||||
соответственно. Пейсмекерные |
|
|||||||
клетки генерируют спонтанный |
|
|||||||
ритм |
|
и |
сопряжены |
с |
|
|||
тормозными |
интернейронами, |
|
||||||
которые |
обеспечивают |
их |
|
|||||
активность |
в |
противофазе. |
|
|||||
Возбуждение |
1 |
пейсмекера |
|
|||||
приводит |
|
к |
|
активации |
|
|||
мотонейрона |
сгибателей |
и |
|
|||||
подавлению |
2 |
пейсмекейкера. |
|
|||||
Тогда же, когда прекращается |
|
|||||||
активность |
1 |
|
пейсмекера, |
|
||||
может |
|
активироваться |
2 |
|
|
|||
пейсмекер, |
|
|
который |
|
|
|||
|
|
Fig. 22.10. A Hypothetical Pattern Generator consisting of two pacemaker cells (1 |
||||||
поддерживает |
заторможенное |
|
||||||
состояние |
1 |
пейсмекера, |
а |
|
and 2) that excite flexor and extensor motoneuron pools, respectively. The |
|||
также |
|
|
|
|
активацию |
pacemaker cells discharge with a spontaneous rhythm and are coupled by |
||
мотонейронов |
разгибателей. |
inhibitory interneurons (black) that ensure that their discharge is out of phase. |
||||||
Две |
|
верхних |
кривых |
When pacemaker 1 fires, the flexor motoneuron is activated and pacemaker 2 is |
||||
представляют |
|
собой |
suppressed. When pacemaker 1 falls silent pacemaker 2 can fire, maintaining the |
|||||
внутриклеточное отведение от |
silence of pacemaker 1 and activating the extensor motoneuron. The upper two |
|||||||
пейсмекера |
1 |
и |
2, нижние |
traces represent intracellular records from pacemakers 1 and 2; the lower traces |
||||
кривые |
|
|
представляют |
represent extracellular recordings from flexor (F) and extensor (E) motoneurons. |
||||
внеклеточное |
отведение |
от |
|
|||||
мотонейронов сгибателей (F) и |
|
|||||||
разгибателей (Е) |
|
|
|
|
||||
трального ритма во многих случаях включает комбинацию пейсмекеров и сетевых механизмов40· 41).
Локомоция
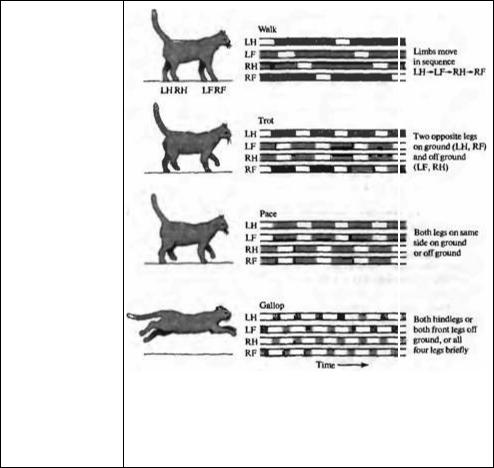
Во время локомоции каждая нога проделывает элементарные шагающие движения, которые состоят из двух компонентов42); (1) качательная фаза, во время которой нога, вытягиваясь назад, сгибается, поднимается над землей и устремляется вперед и вытягивается вновь, чтобы наступить на землю, и (2) устойчивая фаза, когда нога, будучи в контакте с поверхностью, двигается назад по отношению к направлению, выбранному телом. Последовательность движений конечностей является стереотипной реакцией.
На рис. 22.11 показано, что у шагающей · кошки сначала поднимается левая задняя конечность, затем — левая передняя, правая задняя и только потом — правая передняя нога. Если же кошка начинает двигаться быстрее, происходит два типа изменений: (1) генератор движений для каждой конечности укорачивает устойчивую фазу, но вместе с тем усиливает отталкивание от поверхности. Качательная фаза при этом остается относительно постоянной.
(2) Увеличение скорости может также вовлекать изменения походки, зависящей от координации движений конечностей. Движение «рысью» базируется на одновременном поднятии двух конечностей, а галоп предполагает, что от земли могут отрываться все четыре конечности одновременно. Стереотипный характер этих реакций предполагает, что в ЦНС имеется целый ряд двигательных программ, которые «проигрываются» после получения соответствующей команды. Имеется достаточно много свидетельств того, что существуют заранее заложенные, генетически предопределенные двигательные программы, которые появляются спонтанно, независимо от приобретенного опыта43, 44).
Еще в 1911 году Браун45) показал, что в спинном мозге кошки имеются простые нейронные сети, обеспечивающие шагательные движения. Попеременное поднятие и опускание задних конечностей могло наблюдаться у кошки (подвешенной над бегущей дорожкой), у которой спинной мозг был пересечен на уровне грудных сегментов. Дальнейшие эксперименты показали, что у спинальной кошки введение ДОФА (предшественника биогенных аминов — см. главу 9) позволяет сохранять походку, несмотря на ускорение движения дорожки46). Наиболее важное значение этих экспериментов с клинической точки зрения заключается в том, что после пере-
Глава 22. Клеточные механизмы двигательного контроля |
499 |
Рис. 22.11. Четыре различных двигательных паттерна локомоции во время шагания, рыси, иноходи и галопа. Светлые линии показывают время, в течение которого конечность оторвана от земли (качательная фаза, во время которой активны мотонейроны сгибателей). Во время шагания конечности двигаются последовательно, сначала
содной стороны, потом —
сдругой. Во время рыси используется другой паттерн координации конечностей:
противоположные по диагонали конечности поднимаются одновременно. При иноходи ритм меняется вновь: конечности с одной стороны поднимаются одновременно. Горазда более быстрым является галоп, во время которого от поверхности отрываются сначала задние, а потом — передние конечности.
Fig. |
22.11. The |
Stepping |
which flexor motoneurons are active);the light bars show the time a foot is on the ground |
Pattern of a Cat during |
(the stance phase, during which extensor motoneurons are active). During walking, the |
||
walk, trot pace, and gallop, |
legs are moved in sequence, first on one side, then on the other. During a trot a different |
||
four |
different |
gaits of |
pattern of interlimb coordination is used: Diagonally opposite limbs are raised together. |
locomotion. The white bars |
In a pace, the rhythm changes again, the limbs on the same side being raised together. |
||
show the time that a foot is |
Faster still is the gallop, in which the hindlimbs and then the front limbs leave the |
||
off the ground (the swing |
ground. LF = left foreleg, LH = left hindleg, Rf = right foreleg, RH = right hindleg. (After |
||
phase, during |
|
Pearson, 1976.) |
|
сечения спинного мозга биогенные амины помогают адаптироваться к шагательным движениям с возрастающими нагрузками47· 48). Это означает также, что эффективность генетически заложенных двигательных программ может быть повышена за счет практики. В согласии с этим находятся данные о том, что тренировка на бегущей дорожке может обучить движениям при параплегии у человека49). В этих экспериментах запуск движений конечностей происходил в результате контакта с дорожкой, и после определенной практики такого рода происходило улучшение симптоматики. Объяснение этому базируется: по-видимому, на том, что центральные генераторы ритма активируются сенсорным притоком (поскольку дорожка перемещает конечность) и эта рефлекторная реакция усиливается за счет многих повторений.
Взаимодействия сенсорной импульсации и центральных генераторов ритма
Как указывалось ранее, наличие сенсорного притока с периферии не является обязательным условием для генерации ритмической двигательной активности. Так, после отделения нервной системы таракана от сенсорных входов мотонейроны продолжают генерировать импульсы, которые позволяют ему нормально передвигаться50). Аналогично, изолированные сегменты спинного мозга миноги 51, 52), черепахи53· 54), крысы39) сохраняют такие ритмические разряды мотонейронов, которые происходят у интактных животных во время плавания или шагания.
Многие движения, независимо от того, являются ли они ритмическими или нет, могут
500 |
Раздел III. Интегративные механизмы |
совершаться в отсутствие сенсорной импульсации. Например, во время пения птиц последовательные движения мышц происходят так быстро, что просто не оставляют времени для реализации обратной связи от рецепторов: следующая команда из ЦНС поступает еще до того, как происходит анализ последствий предыдущей команды. Как приматы, так и человек могут совершать достаточно тонкие выработанные ранее движения даже в условиях деафферентации конечностей55). Однако в отсутствие обратной связи точность движений начинает страдать по мере усложнения заданий, по-видимому как результат накопления мелких ошибок.
У человека, например, становится нечетким почерк. Кроме того, деафферентация существенно нарушает способность к обучению новым двигательным навыкам, так же, как удаление соматосенсорной коры. После одностороннего удаления полей 1, 2 и 3 (первичная соматосенсорная кора), обучение у приматов незнакомым заданиям на стороне, контралатеральной повреждению, было существенно затруднено56). Интересно, что повреждение коры после выработки двигательных навыков существенно не сказывалось на их выполнении. Таким образом, нельзя говорить о том, что наличие врожденных программ движений в условиях целого организма приводит к полному игнорированию сенсорного потока. Например, легкое касание палочкой стопы кошки во время выполнения качательной фазы шагательных движений приводит к аккуратному перешагиванию через это препятствие. Было показано, что важная информация о состоянии мыщц-удлинителей во время локомоции приходит по Ib афферентам от сухожильных органов Гольджи. Это оказывает немедленное влияние на силу и длительность устойчивой фазы шагательных движений22· 57· 58). Контроль мышечных веретен посредством γ' мотонейронов представляет собой другой способ модуляции двигательной команды. Ко-активация α- и γ-мотонейронов устанавливает длину экстрафузальных мышечных волокон. Если внешнее препятствие мешает нормальному перемещению конечности, усиленный афферентный поток приводит к дополнительному возбуждению мотонейронов. Задача сенсорной импульсации — адаптировать выполнение текущей двигательной команды к изменяющимся условиям функционирования. Взаимодействие сенсорного притока и внутренних генераторов ритма лучше всего представлено на примере простого ритмического акта, каким является дыхание.
Дыхание
От момента перерезки пуповины до последнего дня нашей жизни постоянный приток кислорода и удаление двуокиси углерода обеспечивается непрерывной работой легких. Две антагонистические группы мышц обеспечивают поступление и последующую откачку воздуха из легких. Во время вдоха грудная клетка увеличивается благодаря сокращению межреберных мышц и диафрагмы. В результате этого происходит расширение легких и последующий вход в них воздуха. Другие мышцы грудной клетки и живота также в разной степени принимают участие в акте вдоха и выдоха, в зависимости от положения тела, скорости и глубины дыхания59). Пример дыхательного ритма показан на рис. 22.12. Активность каждой мышцы могла быть зарегистрирована датчиком напряжения или записью электрических сигналов электродом, введенным в саму мышцу — электромиографией. Рис. 22.12 показывает, что сокращения мышц вдоха и выдоха сопровождаются залпами потенциалов действия соответствующим разрядам двигательных единиц. Очевидно, что обе группы мышц сокращаются в противофазе.
Во время дыхания рефлекс растяжения вносит свой вклад в величину вдоха и выдоха, поддерживая возбудимость мотонейронов по мере того, как поочередно внутренние и наружные мышцы подвергаются растяжению. Каждый Ia афферент, генерирующий импульсы с частотой около 100/с, вносит вклад в суммарный ВПСП в гомонимных мотонейронах (то есть мотонейронах, иннервирующих ту же мышцу), а также, через интернейроны, приводит к торможению мышц--антагонистов. В течение всего цикла, благодаря работе фузимоторных волокон (аксонов γ-мотонейронов), афференты поддерживают высокую частоту импульсации даже тогда, когда мышцы активно сокращаются 27· 28) (см. рис. 22.8). Фузимоторные волокна подстраивают длину мышечных веретен к той длине мышцы, которая ожидается во время этой половины цикла. Если какое-то препятствие ограничивает ожидаемое движение, укороченное мышечное веретено будет растягиваться, увеличивая возбудимость соответствующих мотонейронов.
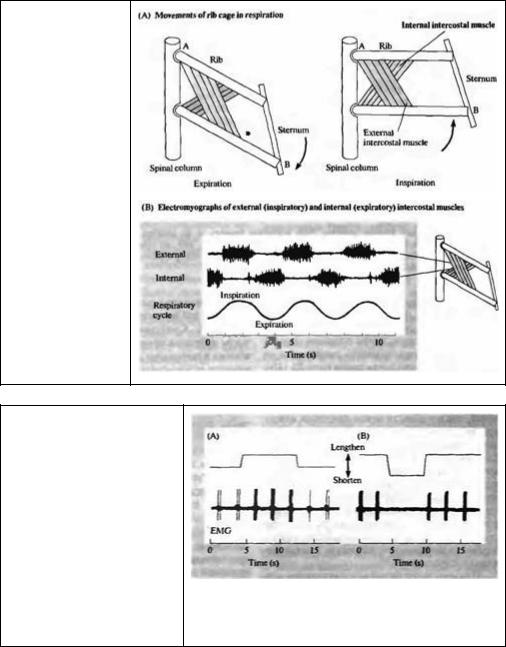
502 |
Раздел III. Интегративные механизмы |
|
|
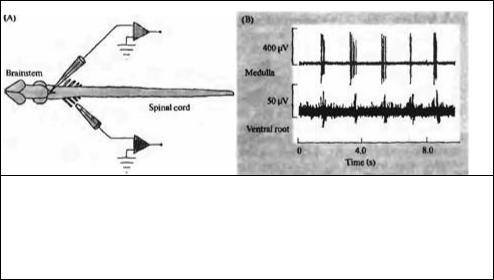
Рис. 22.14. Дыхательный ритм, отводимый от стволовых нейронов изолированной ЦНС новорожденного опоссума (А). (В) Две стволовых клетки генерируют регулярные разряды с межимпульсными интервалами около 2 с. Одновременное отведение от грудных вентральных корешков показывает соответствующие разряды дыхательных мотонейронов.
Fig. 22.14. Respiratory Rhythm Recorded from Brainstem Neurons in an isolated central nervous system from a neonatal opossum (A). (B) Two brainstem cells discharge regularly with an interburst interval of about 2 s. Simultaneous recording from a thoracic ventral root shows the corresponding discharge of respiratory motoneurons.
(Kindly provided by D.J.Zou and 3.G. Nicholls )
существенных изменений паттерна. Хотя единичные ВПСП при активации афферентов и малы, но их суммарный эффект является достаточно мощным, чтобы подавить способность мотонейрона вызывать сокращение во время прекращения растяжения (рис. 22.13В).
Как же генерируется дыхательный ритм? Пересечение продолговатого мозга у кошки приводит к прекращению дыхания59). Это говорит о том, что генератор ритма лежит не в спинном мозге. Как в продолговатом мозге, так и в области моста находятся нейроны, разряжающиеся во время вдоха или выдоха и вызывающие возбуждение и торможение соответствующих мотонейронов. Например, во время вдоха мотонейроны, снабжающие наружные межреберные (инспираторные) мышцы, деполяризуются за счет ВПСП, приходящих из дыхательного центра продолговатого мозга. Инспираторная фаза прекращается
залпом тормозных импульсов, приходящих из центральных нейронов, ассоциированных с выдохом60· 61).
Пример такого ритмически активного нейрона ствола мозга новорожденного опоссума62) показан на рис. 22.14А. Верхняя запись на рис. 22.14В представляет собой внеклеточное отведение от одиночного нейрона в продолговатом мозге. На этой регистрации видны короткие залпы импульсов с периодом около 2 с. На нижней записи отведение от грудных вентральных корешков показывает соответствующие ритмические разряды мотонейронов, снабжающих дыхательные мышцы и следующие с той же частотой, хотя и с небольшой задержкой. Свойства и синаптические контакты нейронов продолговатого мозга, вовлеченных в генерацию дыхательного ритма пока изучены недостаточно. Пейсмекерные осциллирующие нейроны найдены у новорожденных крыс в области вентральной части продолговатого мозга (названной pre-Botzinger complex) и вовлечены в генерацию дыхательного ритма63)--65). В этот процесс вовлечены также реципрокные связи между интернейронами.
Частота и глубина дыхания зависят от уровня СО2 в артериальной крови66). В условиях низкого уровня СО2 частота и глубина дыхания снижаются, и наоборот, дыхание усиливается при повышении уровня СО2. Этот эффект зависит от импульсов, приходящих от хеморецепторов каротидного синуса и аорты, а также от нейронов продолговатого мозга, чувствительных к уровню СО267). Залпы импульсов в индивидуальных нейронах, контролирующих инспираторные и экспираторные мотонейроны, критически зависят от уровня СО2. Вариации концентрации СО2 транслируются в изменения частоты разрядов интернейронов, а от них, соответственно, передаются к мотонейронам. 68) Таким образом, сенсорные входы в стволе мозга играют весьма важную роль в модуляции дыхательного ритма.