

Nikolls_-_Ot_neyrona_k_mozgu
.pdfГлава 24. Денервация и регенерация синаптических связей |
583 |
Рис. 24.5. В мышце крысы блокирование нервной проводимости приводит к появлению новых АХ рецепторов. (А) В нормальной мышце чувствительность к АХ ограничена участком концевой пластинки. (В) После блокады двигательного нерва в течение 7 дней местным анестетиком чувствительность к АХ распространяется по всей поверхности мышечного волокна.
Fig. |
24.5. |
|
New |
ACh |
|
||
Receptors |
appear |
after |
|
||||
block of nerve conduction in |
|
||||||
rat muscle. (A) In the normal |
|
||||||
muscle, |
ACh |
sensitivity |
is |
|
|||
restricted to the end-plate |
|
||||||
region (near the 5 mm |
|
||||||
position). (B) After the nerve |
|
||||||
to the muscle was blocked |
|
||||||
for 7 days by a local |
|
||||||
anesthetic, |
|
the |
ACh |
|
|||
sensitivity is distributed over |
|
||||||
the |
entire |
|
muscle |
fiber |
|
||
surface. |
|
Sensitivity |
|
is |
|
||
expressed |
numerically |
in |
|
||||
millivolts |
of |
depolarization |
ejected from the pipette (Chapter 9). The crosses and bars represent the mean and |
||||
per nanocoulomb of charge |
range of sensitivities of a number (in parentheses) of adjacent muscle fibers. (From |
||||||
|
|
|
|
|
|
|
Lemo and Rosenthat 1972.) |
ся сниженным, что устраняет ингибирование и приводит к повышению экспрессии АХ рецепторов.
Изменение времени полураспада АХ рецептора, происходящее в денервированной мышце, также является результатом снижения мышечной активности42). Скорость деградации рецептора возрастает как в мышцах, парализованных денервацией, так и в тех, которые обездвижены продолжительной аппликацией тетродотоксина. Наоборот, прямая электрическая стимуляция денервированной мышцы восстанавливает число АХ рецепторов в синаптических зонах до исходного нормального уровня. И в этом процессе вход ионов кальция в мышечное волокно через потенциал-активируемые кальциевые каналы играет важную роль43). Снижение скорости деградации АХ рецепторов, индуцированное мышечной активностью, воспроизводится обработкой инактивированных мышц кальциевым ионофором А23187. Напротив, стаби-
лизация АХ рецептора в мембране предупреждается блокаторами кальциевых каналов. Повышение концентрации внутриклеточного цАМФ также замедляет деградацию рецепторов в инактивированных мышцах. Последний факт свидетельствует о том, что приток ионов кальция в мышечное волокно вызывает стабилизацию рецептора через активацию аденилатциклазы и последующие процессы фосфорилирования белка44· 45).
Нервные факторы регуляции синтеза АХ рецептора
Мышечная активность является не единственным фактором, поддерживающим нормальное число и композицию рецепторов в скелетных мышцах. Об этом свидетельствуют эксперименты по частичной денервации мышцы, в которых медленно развивающиеся денервационные изменения наблюдались, несмотря на сохранение мышечной активности. Сделать это заключение помогло то, что волокна

584 |
|
|
|
|
|
|
Раздел IV. Развитие нервной системы |
|
Рис. |
24.6. |
|
Устранение |
|
|
|||
гиперчувствительности |
в |
|
|
|||||
денервированной |
мышце |
|
|
|||||
крысы |
|
|
|
|
прямой |
|
|
|
стимуляцией |
мышечных |
|
|
|||||
волокон. (А) Увеличение |
|
|
||||||
чувствительности |
|
во |
|
|
||||
внесинаптической |
части |
|
|
|||||
мышечного |
|
|
волокна |
|
|
|||
после |
|
|
14 |
дней |
|
|
||
денервации. |
|
|
|
(В) |
|
|
||
Чувствительность |
|
|
|
|
||||
внесинаптического |
|
|
|
|||||
участка |
денервированной |
|
|
|||||
мышцы, |
сохранявшейся в |
|
|
|||||
течение |
7 |
дней |
без |
|
|
|||
стимуляции, |
|
а |
затем |
|
|
|||
стимулированной |
|
с |
|
|
||||
перерывами |
|
|
|
в |
|
|
||
последующие 7 дней. Эта |
|
|
||||||
процедура |
|
устраняла |
|
|
||||
денервационную |
|
|
|
|
||||
гиперчувствительность. (С) |
|
|
||||||
Чувствительность |
к |
АХ |
|
|
||||
двух |
|
стимулированных |
|
|
||||
волокон |
той |
же |
мышцы |
|
|
|||
рядом |
|
|
|
|
|
с |
|
|
денервированными |
|
|
|
|||||
участками |
|
|
концевой |
|
|
|||
пластинки. |
|
|
|
В |
|
|
||
стимулированной |
мышце |
(A) Increased sensitivity in the extrasynaptic portion of a muscle |
fiber after 14 days of |
|||||
высокая |
чувствительность |
denervation. (B) Sensitivity in the extrasynaptic region of a |
muscle that had been |
|||||
к АХ |
|
ограничена этим |
denervated for 7 days without stimulation, and then stimulated |
intermittently for another |
||||
участком. |
|
|
|
|
7 days. This treatment reversed the denervation Supersensitivity. |
(C) ACh sensitivity in |
||
Fig. |
24.6. |
Reversal |
of |
two stimulated fibers of the same muscle near their denervated |
end-plate regions. The |
|||
high sensitivity is confined to this region in the stimulated muscle. (After L0mo and |
||||||||
Supersensitivity |
|
in |
a |
Rosenthal, 1972.) |
||||
denervated |
rat |
muscle |
by |
|
|
|||
direct |
stimulation |
of |
the |
|
|
|||
muscle fibers. |
|
|
|
|
|
|||
длинной портняжной мышцы лягушки иннервируются более чем одним синапсом. Если эта мышца частично денервировалась перерезкой внутримышечных разветвлений нерва, в денервированных участках развивалась типичная гиперчувствительность мышечных волокон к АХ. Однако в целом эти мышечные волокна были активными и сохраняли сократимость по всей длине23). Гибридизация in situ в интактной, денервированной и парализованной токсином мышцах крысы показала, что распределение мРНК, кодирующей синтез АХ рецепторов, независимо от мышечной активности, зависит, по крайней мере, от двух нервных факторов. Один из этих факторов стимулирует экспрессию взрослой ε-субъединицы в ядрах, примыкающих к концевой пластинке, а другой фактор угнетает экспрессию γ-субъединицы и подавляет образование мРНК для других субъединиц39).
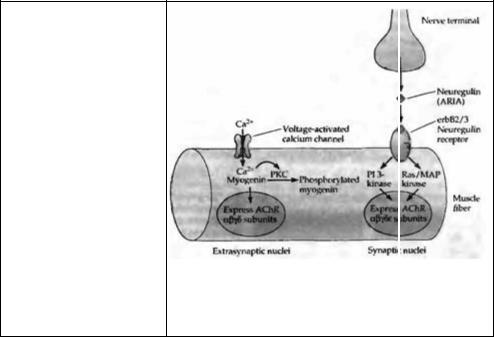
Одним из нервных факторов, регулирующих экспрессию АХ рецептора, является АРМА (ацетилхолиновый рецептор, индуцирующий активность), который был исходно выделен из мозга цыпленка41· 46). АРИА синтезируется и секретируется мотонейронами и стимулирует экспрессию всех пяти субъединиц АХ рецептора, и среди них наиболее заметно — синтез ε- субъединицы (рис. 24.7)47). АРИА принадлежит к семейству белков, называемых нейрорегулинами. Рецепторы для нейрорегулинов относятся к семейству рецепторов тирозинкиназ, родственных с рецептором I типа для эпидермального фактора роста. Установлено, что несколько субъединиц этого рецептора (erbВ2, erbВЗ и erbВ4) аккумулируется в нервно-мышечном соединении. Стимуляция erbВ рецепторов в мышце активирует два внутриклеточных сигнальных каскада, таких как киназу Ras/митогенактивирующего белка (Ras/MAP) и фосфатидилинозитол 3-киназу (PI-киназа), вызывая, таким образом, усиление экспрессии гена АХ рецептора48)--50).
Глава 24. Денервация υ регенерация синаптических связей |
585 |
Рис. 24.7. Контроль синтеза АХ рецепторов ионами кальция и нейрональными факторами. Во внесинаптической части скелетного мышечного волокна позвоночного вход ионов кальция через потенциал-активируемые кальциевые каналы активирует протеинкиназу С (РКС), которая фосфорилирует и инактивиру ет миогенин. В синапсе нейрорегулин ARIA освобождается из нервных окончаний и взаимодействует с erbВ2/3 рецепторами. Это активирует фосфатидилинозитол 3- киназу (PI 3-киназа) и Ras/митогенактивирующую протеинкиназу (Ras/MAP), что ведет к экспрессии
αβ/0γδ/0и |
ε |
-субъединиц |
АХ |
|
рецептора. |
Другие нейрональные |
|
||
сигналы подавляют экспрессию γ- |
|
|||
субъединицы. |
|
|
||
Fig. 24.7. Control of ACh Receptor |
which phosphorylates and inactivates myogenin. At the synapse, the |
|||
Synthesis by Calcium and Neural |
||||
Factors. In extrasynaptic regions of a |
neuregulin ARIA is released from nerve terminât and interacts with erbB2/3 |
|||
vertebrate skeletal muscle fiber, influx |
receptors. This activates phosphatidylinositol 3-kinase (PI 3-kinase) and |
|||
of calcium through voltage-activated |
Ras/mitogenactivated protein (Ras/MAP) kinase pathways, leading to |
|||
calcium |
channels activates |
protein |
expression of AChR a, /9, 7, δ and ε subumts. Other neural signals suppress |
|
kinase С (РКС), |
|
expression of the 7 subunit. |
||
|
|
|||
Распределение рецепторов в нервных клетках после денервации
Эффект денервации на распределение рецепторов в нейронах изучался на автономном ганглии лягушки и цыпленка. В живом сердце лягушки парасимпатические нейроны можно увидеть в прозрачной межпредсердной перегородке, что значительно облегчает аппликацию АХ на отдельные участки клеточной поверхности. Кроме того, клетки не имеют дендритов; синапсы формируются прямо на клеточной соме (рис. 24.8А). Подобно волокнам скелетной мышцы, эти нейроны чувствительны к АХ на участках, располагающихся под пресинаптическими окончаниями5). При оценке распределения АХ рецепторов иммунофлуоресцентной микроскопией было обнаружено, что клетки ганглия имеют приблизительно 30 крупных плотно упакованных кластеров АХ рецепторов, локализующихся в синаптических зонах, и более 100 маленьких внесинаптических кластеров, рассеянных по всей поверхности клетки (рис. 24.8В). Примерно 20 % рецепторов находились во внесинаптических зонах53).
Для изучения эффектов денервации на сердце лягушки перерезали оба блуждающих нерва54· 55). Синаптическая передача между окончаниями блуждающего нерва и клетками ганглия нарушалась уже на второй день после денервации. Параллельно с этим увеличивалась поверхность нейрона, чувствительная к АХ. Через 4—5 дней после денервации аппликация АХ в любой точке клеточной поверхности вызывала деполяризацию мембраны. В других отношениях клетки выглядели неизмененными. Полученный результат был, по-видимому, не следствием изменения числа АХ рецепторов, а скорее был связан с их перераспределением (рис. 24.8С и D)52, 56). Денервация сокращала количество синаптических кластеров на 90 % и вызывала увеличение в 2—3 раза числа малых внесинаптических кластеров, распределенных по всей клеточной поверхности. Денервация приводила также к снижению уровня ацетилхолинэстеразы57). Таким образом, чувствительность к АХ повышалась. Если условия способствовали регенерации исходного нерва, чувствительная к АХ зона вновь ограничивалась синаптическими зонами55).
В других ганглиях цыпленка и лягушки денервация либо имела меньший эффект, либо не оказывала влияния на распределение АХ рецепторов на клеточной поверхности 58)--61). Например, денервация не меняла чувстви-

586 |
Раздел IV. Развитие нервной системы |
Рис. 24.8. (Рис. 24.8В см. на цветной вклейке после с. 640.) Развитие гиперчувствительности в парасимпатических нервных клетках сердца лягушки после денервации. (А) Парасимпатические ганглионарные клетки иннервированы аксонами блуждающего нерва, которые формируют конечные синаптические бутоны, диффузно представленные по всей клеточной поверхности. (В) Парная иммунофлуоресцентная микрография клетки ганглия нормального животного, помеченная антителами к АХ рецептору и к синаптическим везикулам. В синаптических зонах расположены крупные плотно-упакованные кластеры АХ рецепторов; более ста малых кластеров распределены по внесинаптической поверхности. (С, D) Изображение ложнооперированных (С) и денервированных (D) ганглионарных клеток, помеченных антителами к АХ рецептору. Денервация вызывает снижение числа синаптических кластеров и значительный рост небольших внесинаптических кластеров, что приводит к развитию гиперчувствительности.
Fig. 24.8. Development of Supersensitivity in parasympathetic nerve celts in frog heart after denervation. (A) Parasympathetic gangion cells are innervated by axons in the vagus nerve, which form terminal boutons scattered over the cell surface. (B) Stereo-pair immuno-fluorescence micrographs of a ganglion cell in a normal animal, labeled with antibodies to
AChRs and to synaptic vesicles. Large, dense clusters of AChRs are located at synaptic sites; more than a hundred small extrasynaptic clusters are spread over the rest of the cell surface. (C, D) Images of sham-operated (C) and 20day denervated (D) ganglion cells labeled with antibodies to AChRs. Denervation causes a decrease in the number of synaptic clusters and a marked increase in small extrasynaptic clusters, producing supersensitivity. (From Wilson Horch and Sargent, 1996; micrographs kindly provided by P. B. Sargent.)
тельность нейронов симпатического ганглия лягушки к ионофоретически приложенному АХ. Однако, если чувствительность к АХ измерялась в условиях введения его через омывающий раствор, наблюдалось 18-кратное повышение чувствительности. Это различие, по-видимому, было обусловлено снижением активности ацетилхолинэстеразы, ассоциированной с преганглиальными аксонами, которая в интактных условиях предотвращает проникновение АХ к нейронам, располагающимся внутри ганглия. Таким образом, эффекты денервации на распределение рецепторов для нейротрансмиттеров различны в нервных и мышечных клетках. Более того, они различаются и в зависимости от типа нейронов.
Восприимчивость нормальной и денервированной мышцы к новой иннервации
У взрослого млекопитающего и лягушки иннервированные мышечные волокна не допускают иннервации дополнительным нервом62). Так, если приложить перерезанный конец двигательного нерва к иннервированной мышце, в ней не будут формироваться ни новые разветвления, ни дополнительные концевые пластинки. Наоборот, из того же нерва образуются нервные разветвления и происходит реиннервация, если он приложен к денервированной или поврежденной мышце. В отличие от ситуации во время развития нервномышечной системы, когда растущие конусы нерва контактируют с мышечными волокнами беспорядочно, реиннервация обычно происходит в зоне исходной концевой пластинки. Регенерированные аксоны направляются к исходным синаптическим зонам по эндоневрию предшествующего аксона (рис. 24.1) и тянутся отростками к щванновским клеткам, которые покрывали терминали прежнего аксона (глава 8)63). Тем не менее, если перерезанный нерв находится далеко от исходной концевой пластинки или есть факторы, препятствующие росту аксона, то может образоваться новая концевая пластинка. Это значит, что нервные волокна могут расти во взрослом мышечном волокне и образовывать синапсы в участке, который никогда не был иннервирован, с последующим формированием пре- и постсинаптических специализированных структур.
Глава 24. Денервация υ регенерация синаптических связей |
587 |
Гиперчувствительность и формирование синапса
При каких условиях денервированная мышца не отторгает нерв? После того как мышцы крысы стали гиперчувствительными в результате блокирования проведения импульсов в нерве или вследствие аппликации ботулинового токсина, чужие нервы могут образовывать дополнительные синапсы62· 64· 65). После устранения блока каждый из двух нервов может продуцировать синаптические потенциалы и вызывать мышечные сокращения. Наоборот, если денервированная мышца подвергается прямой стимуляции, ее способность воспринимать дополнительную иннервацию теряется вместе с ее гиперчувствительностью. Является ли гиперчувствительность мышцы необходимой предпосылкой для возникновения иннервации? Как описывалось в главе 23, к моменту иннервации мышечные волокна эмбрионов и новорожденных крыс чувствительны к АХ по всей своей поверхности66). Аналогично, мышечные волокна, растущие в клеточной культуре, чувствительны к АХ и поддаются иннервации по всей своей протяженности67). Таким образом, оба феномена, как начальная иннервация, так и реиннервация, происходят тогда, когда мышечные волокна являются гиперчувствительными к медиатору. Тем не менее формирование синапса не зависит напрямую от функции АХ рецептора. В денервированных мышцах крысы и Xenopus реиннервация происходит и тогда, когда АХ рецепторы блокированы α-бунгаротоксином или кураре68· 69). Одним из факторов, обеспечивающих восприимчивость мышцы к иннервации, является MuSK, рецептор тирозинкиназы, через который агрин запускает дифференцировку постсинаптического аппарата (глава 23) 70).
Аксональный рост, индуцированный денервацией
Денервированные мышцы не только поддаются иннервации, но они активно побуждают неповрежденные нервы отращивать новые разветвления (вызывают спраутинг). Например, если мышца частично денервирована, оставшиеся терминали аксонов будут расти и иннервировать денервированные мышечные волокна (рис. 24.9)71). Так же как регуляция синтеза и деградации АХ рецептора, запускающим фактором для роста аксонов является отсутствие мышечной активности.
Спраутинг и гипериннервация происходят тогда, когда мышечная активность устранена блокированием потенциалов действия нерва тетродотоксином72) или если нервно-мышечная передача блокирована ботулиновым токсином или α-бунгаротоксином73· 74). Кроме того, важную роль в регуляции роста конечных разветвлений аксона играют шванновские клетки, покрывающие нервные терминали (глава 8)75).
Молекулярные механизмы, вызывающие рост терминалей аксонов, еше не изучены в достаточных деталях. Однако ясно, что сигналы, контролирующие этот процесс, являются высокоспецифическими. В коже пиявки, например, разрушение отдельных чувствительных или двигательных нейронов с помощью проназы вызывает рост аксонов в денервированную зону, но не любых, а только аксонов клеток той же модальности76)--78). Привлекательным выглядит использование в будущем методов молекулярной биологии для идентификации в одиночных нейронах пиявки генов, специфически регулирующих эти процессы79).
Подобным же образом, когда клетки зубчатой фасции гиппокампа крысы избирательно денервированы удалением с ипсилатеральной стороны энторинальной коры, три оставшихся входа к этим клеткам растут и образуют новые синапсы совершенно определенным и точным образом (рис. 24.10)80). Аксоны противоположной стороны коры головного мозга, которые обычно обеспечивают только редкие единичные входы, растут очень экстенсивно и замешают как структурно, так и функционально ипсилатеральный энториальный вход, который до этого обеспечивал 80 % синаптических контактов. Среди реагирующих на повреждение волокон есть входы от медиальной перегородки; их спраутинг запускается повышением продукции NGF в поврежденном участке.
§3. Регенерация периферической нервной системы позвоночных
Восстановление поврежденных аксонов
Шванновские клетки периферической нервной системы обеспечивают окружение, способствующее регенерации аксона. Стимули-

588 |
Раздел IV. Развитие нервной системы |
Рис. 24.9. Рост нервных окончаний в ответ на частичную денервацию скелетной мышцы млекопитающего. (А) Обычная модель иннервации. (В) Часть волокон денервирована перерезкой нескольких аксонов, иннервирующих мышцу. (С) Аксоны отрастают от терминалей и узлов вдоль претерминальных аксонов неповрежденных мотонейронов, чтобы иннервировать денервированные волокна. (D) После 1 или 2 месяцев отростки, которые контактировали с вакантными концевыми пластинками, сохраняются, в то время как другие отростки исчезают.
Fig. 24.9. Nerve Terminals Sprout in
Response to Partial Denervation of a mammalian skeletal muscle. (A)
Normal pattern of innervation. (B) Some fibers are denervated by cutting a few of the axons innervating the muscle. (C) Axons sprout from the terminals and from nodes along the preterminal axons of undamaged motoneurons to innervate the denervated fibers. (D) After 1 or 2 months, sprouts that have contacted vacant end plates are retained, while other sprouts disappear. (After Brown,
Holland, and Hopkins, 1981.)
рующая рост нейронов активность шванновских клеток является следствием секреции многих трофических факторов, экспрессии на поверхности клеток молекул адгезии и интегринов, а также продукции компонентов внеклеточного матрикса, таких как ламинин81). Например, эксперименты, в которых повреждался седалищный нерв, показали, что в то время, как периферический участок аксона дегенерирует, уцелевшие шванновские клетки в этой области синтезируют в высоких концентрациях два нейротрофических фактора: BDNF (нейротрофический фактор, полученный из мозга) и NGF (рис. 24.11)82· 83). Таким образом, шванновские клетки могут снабжать трофическими факторами BDNF и NGF двигательные, чувствительные и симпатические аксоны, регенерирующие к своим периферическим мишеням. Интересно, что такие «денервированные» шванновские клетки экспрессируют на своей поверхности большое число низкоаффинных рецепторов для NGF и BDNF. Возможно, это поддерживает некоторую тоническую концентрацию этих нейротрофинов, которые служат для выбора правильного направлении для роста регенерирующих аксонов84). После регенерации нервов шванновские клетки прекращают продукцию NGF и BDNF и вновь обеспечивают изоляцию аксонов.
Аполипопротеин E (ApoE), синтезируемый шванновскими клетками и макрофагами, также накапливается в дистальном участке поврежденных периферических нервов и связывается с базальной мембраной шванновских клеток (см. рис. 24.11 )85, 86). АроЕ поддержи-

Глава 24. Денервация υ регенерация синаптических связей |
589 |
||||||||
Рис. 24.10. Спраутинг аксонов в |
|
|
|||||||
ЦНС |
|
млекопитающих. |
(А) |
|
|
||||
Типичная |
гранулярная |
клетка |
|
|
|||||
зубчатой фасции на ее дендритах |
|
|
|||||||
во |
внешнем |
|
молекулярном |
|
|
||||
слое |
|
(OML) |
|
получает |
|
|
|||
множественные |
сина |
птические |
|
|
|||||
входы |
|
от |
ипсилатеральной |
|
|
||||
энторинальной |
коры |
головного |
|
|
|||||
мозга |
и |
единичные |
входы от |
|
|
||||
контралатеральной |
|
коры |
и |
|
|
||||
медиальной |
|
перегородки. |
|
|
|||||
Дендриты |
|
внутреннего |
|
|
|||||
молекулярного |
|
слоя |
(IML) |
|
|
||||
получают входы от медиальной |
|
|
|
||||||
перегородки |
|
|
|
|
и |
|
|
||
коммиссуральных/ассоциативных |
|
|
|||||||
волокон. |
GCL= |
гранулярный |
|
|
|
||||
клеточный слой. (В) Удаление |
|
|
|||||||
ипсилатеральной |
энторинальной |
|
|
||||||
коры |
приводит |
к |
интенсивному |
|
|
||||
росту |
аксонов |
нервных |
клеток |
|
|
||||
энторинальной |
|
коры |
|
с |
|
|
|||
контралатеральной |
стороны |
и |
|
|
|||||
замещению |
ипсилатерального |
|
|
||||||
энторинального |
входа |
в |
OML. |
|
|
||||
Наблюдается |
также |
спраутинг |
Fig. 24.10. Sprouting of Axons in the Mammalian CNS. (A) A typical granule cell in the |
||||||
аксонов, приходящих в DML от |
dentate gyms receives dense synaptic input from the ipsilateral entorhinal cortex, and |
||||||||
медиальной |
|
перегородки. |
sparse input from the contracterai cortex and medial septum on its dendrites in the |
||||||
Коммиссуральные/ассоциативные |
outer molecular layer (OML). Dendrites in the inner molecular layer (IML) receive |
||||||||
волокна в IML расширяют зону |
medial septal and commissural/associational inputs. GCL = granule cell layer. (B) |
||||||||
иннервации. (С, D) Примеры |
Following ablation of the ipsilateral entorhinal cortex, axons from the contralateral |
||||||||
терминальных |
|
разветвлений |
entorhinal cortex sprout extensively and replace the ipsilateral entorhinal input in the |
||||||
аксонов |
от контралатеральной |
OML. There is also some sprouting of axons from the medial septum in the OML. |
|||||||
энторинальной |
коры |
в |
OML |
Commissural/associational fibers in the IML expand their termination zone. (C, D) |
|||||
зубчатой фасции. (С) Норма. (D) |
Examples of terminal arbors of axons from the contralateral entorhinal cortex in the |
||||||||
Два |
месяца |
спустя |
после |
OML of the dentate gyrus. (C) Normal. (D) Two months after lesion of the ipsilateral |
|||||
повреждения |
ипсилатеральной |
entorhinal cortex. (After Délier and Frotscher, 1997.) |
|
||||||
энторинальной коры. |
|
|
|
|
|
||||
вает жизнеспособность нейронов за счет защиты клеток от окислительного повреждения, а также стимуляции роста и адгезии клеток. Тем не менее у нокаутных мышей, лишенных АроЕ, регенерация и ремиелинизация периферических аксонов существенно не меняется87). Однако мыши с отсутствием АроЕ имеют сниженное число немиелинизированных аксонов и, соответственно, сниженную чувствительность к повреждающим тепловым стимулам86).
При повреждении периферического нерва в зоне повреждения активируются факторы, стимулирующие пролиферацию шванновских клеток. Они включают два цитокина: фактор, ингибирующий лейкемию (ФИЛ)88), и Reg-2, являющийся потенциальным митогеном шванновских клеток (см. рис. 24.11)89). Reg-2 специфически экспрессируется в развивающихся и регенерирующих двигательных и чувствительных нейронах, и эта экспрессия усиливается ФИЛ. Антитела к Reg-2 замедляют регенерацию после передавливания седалищного нерва.
Специфичность реиннервации
Для полного восстановления функции после повреждения нервной системы регенерирующие аксоны должны восстановить связи со своими прежними мишенями. Классические эксперименты Лэнгли, подтвержденные позже исследованиями одиночных клеток, показали, что регенерирующие преганглионарные аксоны нейронов автономной нервной системы реиннервируют соответствующие постганглионарные нейроны2). В этом

590 |
|
|
|
|
|
|
|
|
Раздел IV. Развитие нервной системы |
||
Рис. |
24.11. |
Шванновские |
клетки |
|
|
||||||
способствуют регенерации аксонов в |
|
|
|||||||||
периферической |
нервной |
системе |
|
|
|||||||
позвоночных. |
|
После |
аксотомии |
|
|
||||||
дистальный |
участок |
|
аксона |
и |
|
|
|||||
миелиновая |
оболочка |
дегенерируют |
|
|
|||||||
и |
фагоцитируются. |
Пролиферация |
|
|
|||||||
шванновских |
клеток |
стимулируется |
|
|
|||||||
двумя |
цитокинами: |
|
фактором, |
|
|
||||||
ингибирующим лейкемию (LIF), из |
|
|
|||||||||
макрофагов и Reg-2 из окончаний |
|
|
|||||||||
аксонов. |
|
Экспрессия |
|
Reg-2 |
|
|
|||||
усиливается |
LIF. |
Пролиферирующие |
|
|
|||||||
шваннов ские клетки синтезируют два |
|
|
|||||||||
нейротрофических фактора, BDNF и |
|
|
|||||||||
NGF, |
которые |
сохраняются |
на |
|
|
||||||
клеточной |
|
поверхности |
|
их |
|
|
|||||
низкоаффинными |
рецепторами. |
|
|
||||||||
Нейротрофины |
|
поддерживают |
|
|
|||||||
процесс |
регенерации |
аксонов |
и |
|
|
||||||
направляют |
их |
к соответствующим |
After axotomy, the distal portion of the axon and the |
myelin degenerate and |
|||||||
мишеням. Шванновские клетки и |
are phagocytized. Schwann cell proliferation is stimulated by two cytokines: |
||||||||||
макрофаги |
|
синтезируют |
также |
leukemia inhibitory factor (LIF) from macrophages and Reg-2 from axon |
|||||||
аполипопротеин Ε (ΑροΕ), который |
terminals. Expression of Reg-2 is enhanced by LIF. Proliferating Schwann |
||||||||||
способствует выживанию нейронов и |
cells synthesize two neurotrophic factors, BDNF and NGF, which are held |
||||||||||
регенерации их аксонов. |
|
|
|
on the cell surface by low-affinity BDNF/NGF receptors and help sustain |
|||||||
Fig. |
24.11. |
Schwann |
Cells |
Promote |
regenerating axons and guide them to their targets. Schwann cells and |
||||||
macrophages also synthesize apolipoprotein Ε (ΑροΕ), which may help |
|||||||||||
Axon |
Regrowth |
in |
the |
vertebrate |
promote neuron survival and axon regrowth. |
||||||
peripheral nervous system. |
|
|
|
|
|
||||||
механизме имеет значение, по-видимому, сигнал о позиции нейрона, который оказывает влияние на формирование синапса между нейроном и клеткой-мишенью. Этот механизм справедлив как для симпатической, так и двигательной системы. Так, если межреберная мышца, извлеченная из передней части грудной клетки, трансплантируется в область шеи, то она преимущественно реиннервируется аксонами шейного симпатического ствола, расположенными в самых передних участках спинного мозга (рис. 24.12)90). Наоборот, мышцы, трансплантированные из задних областей тела, имеют тенденцию к реиннервации аксонами более дорзальньгх нейронов. Подобная тенденция установлена и для трансплантации симпатических ганглиев различного исходного расположения91). У молодых крыс зависимость от позиции нейрона показана в реиннервации мышц, имеющих множественную иннервацию несколькими сегментами92).
Нервно-мышечные синапсы у новорожденных крыс, головастиков и взрослых тритонов после поперечного пересечения двигательного нерва с высокой точностью воссоединяются с исходной синаптической зоной93) . Одним из механизмов избирательного восстановления синапсов является конкуренция между аксонами. В мышцах саламандры, иннервированных чужим аксоном, чужеродный синапс элиминируется после того, как нормальный нерв восстановил свой синаптический контакт94). У взрослых млекопитающих чувствительные, двигательные и постганглионарные аксоны показывают более низкую избирательность по отношению к их исходным мишеням. Чужие нервы при иннервации мышечных волокон могут быть столь же эффективными, как и оригинальные нервные волокна93). Более того, чужой нерв может даже вытеснить исходный аксон в интактных мышцах взрослой крысы95). Избирательная регенерация у взрослых млекопитающих возможна, если периферический нерв был не перерезан, а передавлен так, чтобы сохранились эндоневрий и базальная мембрана шванновских клеток, окружавших интактные аксоны (см. рис. 24.1)93). При таких условиях регенерирующие аксоны направляются вновь к своим исходным мишеням. Если эндоневрий был разрушен, например при перерезке нерва, регенерация происходит беспорядочно и аксоны часто создают синапсы с неподходящими мишенями.

Глава 24. Денервация и регенерация синаптических связей |
591 |
Рис. 24.12. Селективная реиннервация мышц, основанная на сегментарном происхождении. (А) Межреберная мышца из грудного сегмента ТЗ, Т4 или Т5 была трансплантирована на шею взрослой крысы и после удаления верхнего шейного ганглия была реиннервирована преганглионарными аксонами шейного симпатического ствола. Сегментарное происхождение входов было определено регистрацией активности мышц при стимуляции отдельных вентральных корешков, которые соединяют преганглиальные аксоны с нервным стволом. (В) Распределение входов к трансплантированным ТЗ, Т4 и Т5 мышцам. Передние мышцы склонны к реиннервации передними преганглиальными аксонами.
Fig. 24.12. Selective Reinnervation of Muscles Based on Segmentai Origin. (A) An intercostal muscle from thoracic segment T3, T4, or T5 was transplanted to the neck of an adult rat and reinnervated by preganglionic autonomie axons in the cervical sympathetic trunk, after removal of the superior cervical ganglion. The segmentai origin of the inputs was determined by recording from the muscle and stimulating individual ventral roots that supply preganglionic axons to the trunk. (B) Distribution of inputs to transplanted T3, T4, and T5 muscles. More anterior muscles tend to be innervated by more anterior preganglionic axons. (After Wigston and Sanes, 1985.)
Свойства нерва и мышцы после образования синапса чужим нервом
Первые наблюдения о последствиях формирования синапсов чужим нервом относятся к 1904 году, когда Лэнгли и Андерсон провели выдающееся исследование, показавшее, что мышцы кошки могут иннервироваться преганглионарными холинергическими симпатическими волокнами96), которые в норме создают синапсы в автономном ганглии (глава 16). Было также показано формирование подобных синапсов между автономными нервами лягушки и скелетными мышцами крысы (рис. 24.12)90· 97). Многие свойства нерва и мышцы в таких экспериментах оставались без изменений, несмотря на необычную иннервацию.
В других экспериментах было показано, что свойства мышц с чужеродной иннервацией заметно изменяются. Ярким примером являются медленные скелетные мышечные волокна лягушки: они иннервированы диффузно, имеют характерные детали строения и не генерируют регенеративных потенциалов действия или быстрых мышечных сокращений98). После денервации медленные волокна могут быть реиннервированы нервами, которые в норме иннервируют быстрые мышцы. В этих условиях медленные волокна становятся способными генерировать потенциалы действия и осуществлять быстрое мышечное сокращение99). Экклс и коллеги после перерезки меняли местами нервы, иннервирующие два типа мышц котят и крыс, различающихся по скорости сокращения. Оба этих типа мышечных волокон генерируют распространяющиеся потенциалы действия и называются медленно- и быстро-сокращающимися волокнами, соответственно. После реиннервации чужим нервом медленно-сокращающиеся мышцы становились быстрее, а быстро-сокращающиеся — медленнее100). Главным фактором трансформации являлся паттерн импульсов в нерве и результирующие мышечные сокращения; мотонейроны, иннервирующие медленно- и быстро-сокращающиеся мышечные волокна, возбуждаются с различной частотой101).
592 |
Раздел IV. Развитие нервной системы |
§ 4. Роль базальной мембраны в регенерации нервно- мышечных синапсов
Структурой, играющей ключевую роль в регенерации нервно-мышечных синапсов, является синаптическая базальная мембрана, которая находится между нервным окончанием и мышечной мембраной. Синаптическая базальная мембрана представляет собой интенсивно окрашиваемое внеклеточное вещество, состоящее из протеогликанов и гликопротеинов. Как показано на рис. 24.1 3А, базальная мембрана окружает мышцу, нервное окончание и шванновскую клетку и погружается в складки постсинаптической мембраны.
МакМахан и его коллеги провели серию элегантных исследований роли синаптической базальной мембраны в дифференцировке нерва и мышцы102)--105). Успех их работы базировался на использовании очень удобной экспериментальной модели тонкой кожно--грудной мышцы лягушки, в которой положение концевых пластинок легко различимо в живой мышце. На первом этапе клетки в определенном участке мышцы локально разрушали либо перерезкой нерва и мышечных волокон, либо повторным приложением металлической пластины, охлажденной жидким азотом (рис. 24.13В). В течение нескольких дней часть мышечных волокон в участке повреждения дегенерировала вместе с нервными окончаниями и была фагоцитирована, но базальная мембрана оставалась при этом интактной (рис. 24.13C). Место исходного нервно-мышечного соединения могло быть легко распознано по характерной морфологии базальной мембраны мышцы и шванновских клеток, остающихся в зоне синапса, а также по наличию холинэстеразы, сохраняющейся на синаптической базальной мембране и в синаптических складках.
Через две недели после повреждения в зоне синаптической базальной мембраны формировались новые мышечные волокна, контактирующие с регенерирующими аксональными окончаниями. Стимуляция нервов вызывала сокращения вновь образованных мышечных волокон. Почти все регенерированные синапсы локализовались точно на исходных синаптических зонах, что выявлялось после окраски мышцы на присутствие холинэстеразы. Таким образом, был сделан вывод о наличии сигналов, ассоциированных с синаптической базальной мембраной, которые определяют успех регенерации синапсов.
Синаптическая базальная мембрана и формирование синаптической специализации
Для дальнейшего исследования природы сигналов, ассоциированных с синаптической базальной мембраной, мышцы повреждали, нерв раздавливали, а регенерацию мышечного волокна предотвращали рентгеновским облучением. Регенерирующие аксоны росли к исходным синаптическим зонам, что выявлялось окраской на холинэстеразу, и формировали активные зоны для освобождения медиатора точно напротив участков базальной мембраны, связанных со вторичными синаптическими складками, — и все это происходило в отсутствие клеточных элементов постсинаптической мишени (рис.24.13D).
В параллельной серии экспериментов МакМахан и его коллеги продемонстрировали, что синаптическая базальная мембрана в регенерирующих мышечных волокнах содержит факторы, запускающие дифференцировку постсинаптической мембраны. Мышцы повреждались, как было описано ранее, а реиннервация предотвращалась удалением большого сегмента нерва. При регенерации новые мышечные волокна образовывали вторичные складки и кластеры АХ рецепторов и ацетилхолинэстеразы точно в зоне контакта с исходной синаптической базальной мембраной (рис. 24.14). Таким образом, сигналы, ассоцированные с синаптической базальной мембраной, при регенерации могут инициировать формирование синаптических специализаций как в мышечных волокнах, так и в нервных окончаниях.
Идентификация агрина
Для идентификации сигнала, связанного с базальной мембраной и инициирующего постсннаптическую дифференцировку, МакМахан и его коллеги использовали морского ската Torpedo califomica. Из электрических органов этого животного, родственных скелетной мышце, они приготовили экстракты, содержащие базальные мембраны. Добавленные к культуре мышечных волокон, экстракты симулировали эффекты синаптической базальной мембраны на регенерацию мышечных волокон, а именно индуцировали формирование