

Nikolls_-_Ot_neyrona_k_mozgu
.pdfГлава 22. Клеточные механизмы двигательного контроля |
503 |
§4. Организация двигательных путей
Организация спинальных мотонейронов
Рис. 22.15 показывает порядок расположения мотонейронов в поперечном срезе спинного мозга на уровне шейных сегментов. Мотонейроны мышц-разгибателей располагаются впереди по отношению к мотонейронам сгибателей. Мотонейроны, снабжающие туловище и проксимальные конечности, располагаются впереди и более центрально. Для них характерна устойчивая активность, поддерживающая позу и осанку. Латеральные мотонейроны иннервируютдистальные мышцы, для которых характерны фазные движения69). Активность медиальных мотонейронов координируется длинными спинальными интернейронами, которые посылают свои отростки на несколько сегментов спинного мозга. Короткие спинальные интернейроны представлены в основном в шейном и поясничном расширениях спинного мозга, где они координируют латеральные мотонейроны.
Супраспинальный контроль мотонейронов
Главный нисходящий тракт к мотонейронам начинается в коре больших полушарий и в стволе мозга (рис. 22.16, см. также приложение С). В соответствии с тем, какой части передних корешков серого вещества они достигают, эти пути разделяются на две группы: латеральные и медиальные 70). Два главных латеральных тракта представляют собой латеральный кортикоспинальный тракт, начинающийся в коре мозга и руброспинальный тракт,
начинающийся в красном ядре среднего мозга. Медиальные пути включают вентральный кортикоспинальный тракт, латеральный и медиальный вестибулоспинальные тракты,
ретикулоспинальные тракты, идущие от моста и продолговатого мозга, а также
тектоспинальный тракт.
Читателя, не близко знакомого с данной темой, эти термины могут привести в замешательство. К счастью, здесь есть достаточно простые правила. Тракты называются по участкам мозга, с которого они начинаются и на котором заканчиваются. Вдобавок в названии есть указание на то, в какой части спинного мозга они проходят (вентральный, медиальный). Ряд терминов, указывающий локализацию трактов, указан в приложении С.
Латеральные двигательные пути
Латеральный кортикоспинальный тракт начинается в моторной и премоторной области коры больших полушарий мозга впереди от центральной извилины (поля 4 и 6 по Бродману, см. приложение С), а также от небольшой полоски в постцентральной области (поле 3, рис. 22.17). Волокна направляются вниз через внутреннюю капсулу и ножки мозга к пирамидам продолговатого мозга, после чего они пересекают срединную линию и продолжаются дальше в составе латеральных нисходящих трактов спинного мозга. Волокна латерального кортикоспинального тракта оканчиваются преимущественно на интернейронах и мотонейронах латеральной группы, которые контролируют дистальную группу мышц, отвечающую за тонкие движения пальцев71). Многие окончания этого тракта переключаются прямо на мотонейроны, контролирующие мышцы пальцев72· 73). У человека и других приматов перерезка латерального кортикоспинального тракта приводит прежде всего к потере способности независимого движения пальцами и,
Рис. 22.15. Организация мотонейронов, снабжающих верхние конечности, показанная в поперечном сечении спинного мозга в шейной области. Мышцы плечевого пояса и рук представлены более медиально, тогда как мышцы кистей — более латерально. Мотонейроны мышц разгибателей локализуются на самом краю серого вещества; мотонейроны сгибателей — более центрально.
Fig. 22.15. Organization of Motoneurons supplying the upper extremities, shown in a transverse section of the spinal cord in the cervical region. Muscles of the shoulder and arm are represented most medially, those of the hand most laterally. Extensor motoneurons are located nearest the margin of the grey matter; flexor motoneurons are more central.
504 |
Раздел III. Интегративные механизмы |
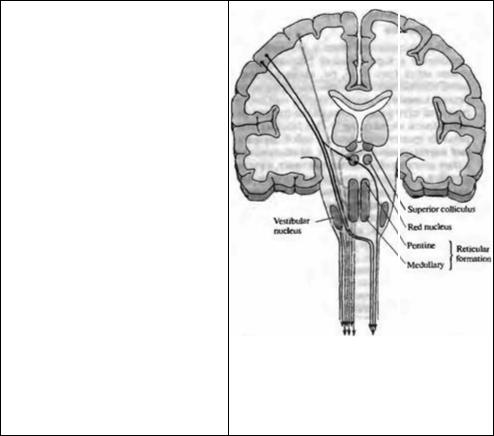
Рис. 22.16. Главные двигательные пути в ЦНС позвоночных, снабжающие латеральные мотонейроны (черные) и медиальные мотонейроны (серые), показанные схематически на срезах полушарий головного мозга, продолжающихся в продольные срезы ствола мозга и спинного мозга. Клетки в первичной двигательной зоне посылают аксоны на контралатеральную сторону спинного мозга, формируя латеральный кортикоспинальный тракт с коллатералями к красному ядру. Аксоны клеток красного ядра пересекают срединную линию и спускаются в составе руброспинального тракта. Эти тракты переключаются моно- и полисинаптически в основном на латеральных мотонейронах. Некоторые кортикальные волокна спускаются без пересечения, формируя передний кортикоспинальный тракт с коллатералями к ядрам ствола. Аксиальная мускулатура снабжается преимущественно моторными зонами ствола через: ретикулоспинальныи тракт, начинающийся с ретикулярной формации ствола мозга и моста; ве стибулоспинальный тракт, начинающийся в вестибулярных ядрах; тектоспинальный тракт, начинающийся в верхнем четверохолмии.
Fig. 22.16. Major Motor Pathways in the vertebrate |
|
|||||
central |
nervous |
system |
supplying |
lateral |
|
|
motoneurons (black) and medial motoneurons (grey), |
|
|||||
shown schematically on a coronal section of the |
|
|||||
cerebral hemispheres, continuing to a longitudinal |
motoneurons (i. e , those supplying distal musculature). Some |
|||||
section of the brainstem and spinal cord. Cells in the |
||||||
cortical fibers descend without crossing to form the ventral |
||||||
primary motor area of the cerebral cortex send axons |
||||||
to the contracterai spinal cord to form the lateral |
corticospinal tract, supplying collaterals to brainstem nuclei. |
|||||
corticospmal tract, with collateral connections to the |
The axial musculature is supplied predominantly by the motor |
|||||
red nucleus. Axons from cells in the red nucleus |
regions of the brainstem through the reticulospinal tracts, |
|||||
cross the midline and descend in the rubrospinal |
originating in the pontine and medullary reticular formations; |
|||||
tract. |
These tracts |
supply |
monosynaptic |
and |
the vestibulospinal tracts, originating in the vestibular nuclei; |
|
polysynaptic innervation largely to lateral |
|
and the tertospinal tract, originating in the superior colliculus. |
||||
|
|
|||||
как следствие, к нарушению тонких, точных движений кисти74· 75).
Руброспинальный тракт начинается в красном ядре (рис. 22.16) и пересекает срединную линию еще до того, как начинает спускаться в спинной мозг. Он заканчивается на интернейронах и, частично, на мотонейронах76), ассоциированных с латеральной двигательной системой. Клетки в красном ядре расположены соматотопически и получают возбуждающие входы от моторной коры и мозжечка. Точная роль руброспинального тракта не известна. По-видимому, он дублирует многие функции кортикоспинального тракта и представляет, таким образом, параллельный ему путь. У приматов повреждение этого тракта вызывает лишь незначительные изменения. Однако после повреждения как руброспинального, так и кортикоспинального трактов координированные движения рук и ног значительно нарушаются77· 78).
Медиальные двигательные пути
За исключением небольшого компонента от вентрального кортикоспинального тракта, медиальные пути начинаются преимущественно в стволе мозга (рис. 22.16) и посылают свои волокна к медиальным мотонейронам, иннервирующим проксимальную группу мышц. Нейроны, начинающие латеральный вестибулоспинальный тракт, лежат (как показывает название) в латеральном вестибулярном ядре. Каждое вестибулярное ядро получает входы от ипсилатерального вестибулярного аппарата (см. главу 17). Этот тракт спускается без пересечений в спинной мозг к медиальной группе мотонейронов, снабжающих позные мышцы. Здесь окончания формируют моносинаптические возбуждающие входы на мотонейронах мышц-разгибателей и, переключаемые через интернейроны, тормозные входы на сгибатели. Функция латерального вестибулоспинального тракта заключается в регуляции поло-
Глава 22. Клеточные механизмы двигательного контроля |
505 |
|||||||
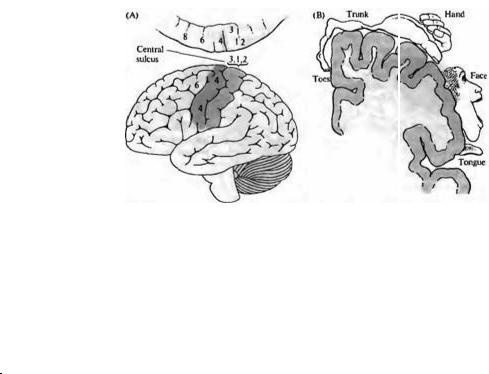
Рис. |
22.17. |
Двигательное |
|
|
||||
представительство в коре. |
|
|
||||||
(А) |
Боковой |
|
вид |
|
|
|||
поверхности |
|
|
коры |
|
|
|||
больших |
|
|
полушарий. |
|
|
|||
Двигательная |
активность |
|
|
|||||
связана |
с |
|
активацией |
|
|
|||
нейронов в поле 4 коры |
|
|
||||||
больших |
|
|
полушарий, |
|
|
|||
включая |
|
|
клетки, |
|
|
|||
формирующие |
|
|
|
|
|
|||
кортикоспинальный |
тракт. |
|
|
|||||
Это |
первичная моторная |
|
|
|||||
зона |
М1. |
|
Моторная |
|
|
|||
система |
включает |
также |
|
|
||||
поле |
6 |
|
(премоторная |
|
|
|||
область), |
|
|
|
|
|
|
|
|
распространяющееся |
на |
|
|
|||||
медиальную |
|
|
|
|
Очень большая область связана с кистями рук, что отражает большое число |
|||
поверхность |
|
полушарий. |
||||||
(В) |
Срезы |
мозга |
через |
нейронов, отвечающих за контроль движений пальцев рук. |
||||
полушария |
впереди |
от |
Fig. 22.17. Motor Representation on the Cerebral Cortex. (A) Lateral view of the surface |
|||||
центральной |
|
борозды. |
||||||
Мускулатура |
|
|
|
|
of the cerebral cortex. Motor activity is associated with activation of cells in area 4 of the |
|||
|
|
|
|
cerebral cortex, including the cells of origin of the corticospinal tract. This is the primary |
||||
человеческого |
|
тела |
||||||
представлена |
|
|
в |
motor area M,. The motor system also includes area 6 (premotor area), extending onto |
||||
последовательном, |
|
но |
the medial surface of the hemisphere. (B) Coronal section through the cerebral |
|||||
непропорциональном |
|
hemisphere anterior to the central sulcus. The |
musculature of the human body is |
|||||
|
represented in an orderly, but distorted, fashion, with the leg and foot on the medial |
|||||||
порядке, |
с |
|
ногами |
и |
||||
ступнями, |
|
|
|
|
|
surface of the hemisphere, and the head most lateral. The very large area devoted to the |
||
|
|
|
|
|
hand is indicative of the number of neurons involved in control of manipulations by the |
|||
представленными |
|
на |
||||||
медиальной |
поверхности |
digits. |
|
|||||
и |
головой |
— |
|
в |
|
|
||
латеральной области. |
|
|
|
|||||
жения тела в пространстве и формировании разгибательного (препятствующего силе тяжести) тонуса. Ретикулоспинальный тракт моста спускается по своей стороне без пересечений и переключается на интернейронах, которые, в свою очередь, обеспечивают двухстороннее возбуждение медиальных мотонейронов разгибателей. Ретикулоспинальный тракт продолговатого мозга спускается с двух сторон для торможения мотонейронов, контролирующих проксимальные конечности.
Два других медиальных тракта ствола мозга заканчиваются на уровне шейных и верхних грудных сегментов спинного мозга и связаны с контролем мыши верхней части туловища, положением рук и особенно головы. Медиальный вестибулоспинальный тракт начинается с нейронов медиального вестибулоспинального ядра, которое, в свою очередь, получает импульсы от полукружных каналов и рецепторов растяжения шеи7). Этот тракт спускается по своей стороне до уровня середины грудной клетки и отвечает за поддержание положения шеи и верхних конечностей во время углового ускорения. Тектоспинальный тракт начинается в верхних ядрах четверохолмия и пересекает срединную линию еще до того, как начинает спускаться к верхним шейным сегментам. Этот тракт отвечает за перемещение головы и глаз к зрительным и слуховым раздражителям.
Таким образом, нисходящие пути, происходящие от коры больших полушарий и красного ядра, снабжают латеральные мотонейроны и выполняют важную функцию по организации движений малых мышечных групп, и особенно дистальных мышц конечностей. В частности, кортикоспинальный тракт важен для движений пальцев. Двигательные пути, исходящие от ствола мозга, наоборот, управляют большими группами проксимальных мышц, имеющих значение для положения тела в пространстве, позиции рук и головы. Эти пути находятся под ключевым контролем со стороны вестибулярного аппарата.
§ 5. Двигательная кора и выполнение произвольных движений
Кортикоспинальный тракт начинается в первичной и вторичной моторных зонах, локализованных в прецентральной извилине, а также от первичной и вторичной соматосенсорной коры в постцентральной извилине (рис. 22.17 и 22.18)80). Нейроны, с которых начинается этот тракт, расположены
506 |
|
|
|
|
|
|
Раздел III. Интегративные механизмы |
|||
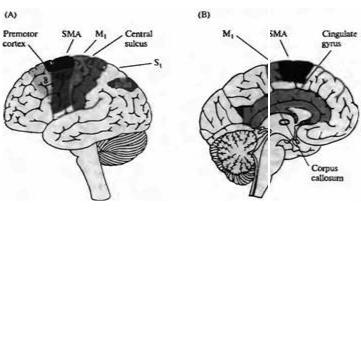
Рис. 22.18. |
Ассоциативная |
|
|
|
|
|||||
двигательная кора. (А) |
|
|
|
|
||||||
Первичная |
|
|
|
и |
|
|
|
|
||
ассоциативная |
|
области |
|
|
|
|
||||
двигательной |
|
коры |
|
|
|
|
||||
располагаются |
|
впереди |
|
|
|
|
||||
центральной |
борозды |
в |
|
|
|
|
||||
поле |
4 |
по |
Бродману |
|
|
|
|
|||
(первичная |
|
двигательная |
|
|
|
|
||||
кора) |
и |
|
поле |
6 |
|
|
|
|
||
(премоторная |
кора |
и |
|
|
|
|
||||
дополнительная |
|
|
|
|
|
|
||||
двигательная зона, SMA). |
|
|
|
|
||||||
Фронтальные |
зрительные |
|
|
|
|
|||||
области найдены в поле 8. |
|
|
|
|
||||||
Первичная |
соматическая |
|
|
|
|
|||||
кора, S1 (поля 3, 1 и 2) и |
|
|
|
|
||||||
особенно |
ассоциативная |
|
|
|
|
|||||
соматосенсорная |
кора |
|
Fig. 22.18. The Association Motor Cortices. (A) The primary |
and association motor |
|
|||||
(поля 5 и 7) генерируют |
|
|
||||||||
cortices lie anterior to the central sulcus in Brodmann's areas 4 |
(primary motor cortex) |
|||||||||
команды, используемые |
в |
|
||||||||
планировании |
движений. |
|
and 6 (premotor cortex and supplemental motor area, or SMA). |
Frontal eye fields are |
||||||
(В) |
|
|
Медиальная |
|
found in area 8. Primary somatic cortex, S1 (areas 3, 1, and 2), and especially |
|||||
поверхность |
полушарий |
|
association somatosensoty cortex (areas 5 and 7), generates commands used in |
|||||||
головного |
мозга. |
Поясное |
|
motor planning. (B) Medial surface of the cerebral hemisphere. The cingulate motor |
||||||
двигательное |
поле лежит |
|
area lies between the cingulate gyrus proper and the medial extension of the primary |
|||||||
между |
собственно |
поясной |
|
motor and supplementary, motor areas |
||||||
извилиной |
и |
медиальной |
|
|
|
|
||||
частью |
|
первичной |
и |
|
|
|
|
|||
дополнительных |
|
|
|
|
|
|
||||
двигательных зон. |
|
|
|
|
|
|
||||
в определенном порядке, образуя соматотопический рисунок в первичной моторной коре — M1 (рис. 22.17В). Этот факт может быть установлен при прямой электрической стимуляции небольших участков этой области, что приводит к сокращению соответствующих мышц. Кроме того, есть соматотопические проекции от соматосенсорной коры прямо к соответствующим колонкам нейронов в зоне М1 81· 82).
Соматотопическая картина двигательной коры была впервые установлена Фритшем и Хитцигом в 1870 году при стимуляции у животных разных участков коры83). Соматотопическое представление тела у человека было впервые показано Пенфилдом с коллегами во время нейрохирургических операций84). Локальная стимуляция участков коры короткими импульсами приводила к движениям мышц в строго определенной области тела. Так же как и в соматосенсорной коре, в моторной зоне тело представлено не пропорционально, с гораздо большим представительством лица и рук по сравнению с туловищем. Неинвазивные современные методы, такие как магнитно-резонансная томография, подтвердили такую организацию моторной коры85). Этот метод в сочетании с транскраниальной стимуляцией, а также ряд других методов позволили установить, что картина представительства мышц в моторной коре достаточна пластична и может быть изменена после периферических повреждений86). Более того, корковое представительство может быть изменено даже после тренировок, приводящих к появлению новых двигательных навыков87)--89). Предполагается, что перераспределение удельных весов синаптических контактов в зоне М1 может представлять собой основу обучения таким навыкам.90)
Ассоциативная двигательная кора
Вторичная или ассоциативная моторная кора состоит из премоторной зоны (поле 6 по Brodmann), которая лежит впереди и несколько латеральнее зоны М1, и дополнительной моторной зоны, также находящейся впереди от М1 (рис. 22.18). Обе эти зоны организованы соматотопически91· 92) (хотя и не так четко как M1) и получают входы от сенсорных ассоциативных зон (задние париетальные поля 5 и 7). Премоторный комплекс находится под сильным влиянием со стороны мозжечка, а дополнительная моторная зона связана с базальными ганглиями. Движения, вызванные электрической стимуляцией премоторной и дополнительной ассоциативных зон являются достаточно сложными и часто вовлекают мышцы с обеих сторон тела.

Глава 22. Клеточные механизмы двигательного контроля |
507 |
Рис. 22.19. (См. цветную вклейку после с. 640.) Функциональная ядерно-магнитная томография двигательной коры человека, активированной во время письма. Каждое изображение коры кодировано в цвете, показывающем области, в которых повышена активность. Четыре верхних изображения показывают активность, когда палец ноги использовался для изображения зигзага (левая пара) или имени субъекта (правая пара). Четыре нижних изображения построены по тому же принципу, но для письма с использованием пальцев руки. Процедура вычитания изображений, результат которого показан на двух левых рисунках, позволила вычислить области, активируемые во время подписывания своего имени, независимо от того, пальцы ног или рук были для этого использованы. Активной оказалась премоторная кора в области представительства кисти, так же как ассоциативная соматосенсорная кора и область латерального мозжечка
Fig. 22.19. Functional MRI of Human Motor Cortex activated during signature writing. Each cortical image is color-coded to indicate areas in which activity is higher than at rest. The upper four images show activity when the toe was used to write a zigzag (left pair) or the subject's name (right pair). The bottom four images are arranged similarly for writing with the finger. Subtractive analysis was used to construct the two images on the right which show areas that were activated during writing of the signature, whether finger or toe was used. The premotor cortex in the region of the hand representation was active, as well as the somatosensory association cortex and a region of the lateral cerebellum. (From Rijntjes et al., 1999.)
Связанная с движениями активность наблюдалась в обеих этих зонах при анализе мозга человека методом позитронно-эмиссионной томографии93). Обе зоны дают соматотопические проекции в первичную моторную зону. Предполагается, что вместе с сенсорными полями, такими как задняя париетальная область, премоторная и дополнительная моторная кора принимают участие в планировании или программировании двигательных актов94)--96).
Яркий пример программирования движений премоторной корой заключается в разных способах, которыми человек может поставить свою подпись. Обычно эта процедура у большинства людей достигает совершенства и выполняется доминирующей рукой. Однако эта двигательная программа может быть перенесена на совершенно другую группу мышц. Так, Эдриан97) показал, что после того, как вы научились ставить свою подпись, вы можете сделать это даже, если держите карандаш между пальцами ног! Это значит, что в двигательной системе должна быть размещена программа выполнения подписи, вне связи с какой-то определенной группой мышц. Методом магнитно-резонансной томографии была проанализирована активность мозга во время выполнения подписи пальцами рук или ног. Эта активность сравнивалась с выполнением случайной двигательной программы (нарисовать зигзаг)98). Оказалось, что при выполнении подписи была активирована премоторная кора в участке, контролирующем кисть, причем независимо от того, исполнялась ли подпись действительно рукой или ногой (рис. 22.19). Итак, выученная двигательная программа хранится в ассоциативной коре в зоне ответственности за тренированную конечность, но эта программа может быть выполнена и совсем другой конечностью.
Еще одна область с особыми двигательными функциями локализуется в области поясной извилины под дополнительной моторной зоной99). Поясной комплекс лежит выше мозолистого тела (рис. 22.18) и представляет собой часть лимбической системы, контролирующей поведенческие акты100). Нейрональная активность в этой области связана с движениями, подкрепляемыми вознаграждением101).
Активность кортикальных нейронов
Как активность нейронов в моторной коре связана с началом и выполнением движений? Способны ли нейроны в Μ1-зоне прямо управлять активностью отдельных мышц, силой сокращения специфических мышечных групп, величиной смещения относительно сустава, а также влиять на направление движения? Такие вопросы поставил перед собой Эвартс102· 103), который отводил активность пирамидных клеток в моторной коре обезьяны во время выполнения заученных движений кисти (рис. 22.20). С помощью нагрузки, уравновешивающей сгибание или разгибание кисти, Эвартс смог отделить силу, необходимую для выполнения движения, от его направления. Было обнаружено, что часть кортикальных нейронов активна во время сгибания, а другие — во время разгибания, а интенсивность их разрядов пропорциональна силе, с которой выполняется движение104). Это поведение существенно не отличалось от реакции спинальных мотонейронов, с которыми кортикальные нейроны были связаны. Последующие эксперименты показали, что конкретная реакция (участие в сгибании или разгибании) является характерной для
508 |
Раздел III. Интегративные механизмы |
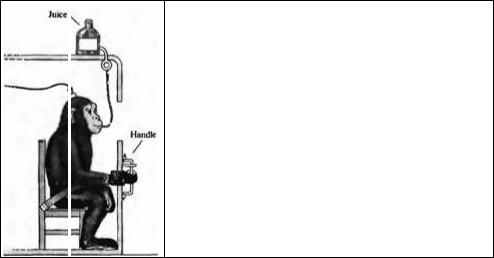
Рис. 22.20. Схема эксперимента по регистрации активности нейронов во время движений кисти. Обезьяна, предварительно обученная передвигать рукоятку в определенное положение, сидит на стуле с манжетами, одетыми на предплечье. Обезьяна отклоняет рукоятку влево или вправо, сгибая или разгибая запястье. Система грузов или вращающих двигателей используется для уравновешения сгибания или разгибания. Для визуального контроля движений положение рукоятки изображается на дисплее. В случае успешного достижения необходимой позиции, обезьяна получает вознаграждение в виде фруктового сока. Активность одной нейромоторной единицы регистрируется микроэлектродом, введенным в соответствующую зону мозга с помощью микроманипулятора, фиксированного к черепу.
Fig. 22.20. Experimental Arrangement for Recording Cellular Activity related to wrist movement. A monkey, previously trained to move a handle to a designated position, is seated in the chair with its forearm placed in a cuff. The monkey deflects a handle to the left or right between stops, by flexion or extension of the wrist. A system of weights, or a torque motor (not shown), is used to load the handle to oppose either flexion or extension. For visually guided movements, the handle position is indicated on a display screen. When the monkey places the handle in the designated position, it receives a reward of fruit juice. Single-unit activity is recorded with a microelectrode positioned in an appropriate area of the brain, by means of a microdrive fixed to the skulL
определенных кортикоспинальных нейронов, прямо оканчивающихся на мотонейронах72). Другие классы нервных клеток демонстрируют более сложное поведение, зависящее от исходной нагрузки и начальной позиции конечности105).
Активность корковых нейронов, связанная с направлением движения
Для того, чтобы дотянуться и схватить определенный объект, нервная система должна совершить ряд вычислений. Зрительный анализатор определяет объект и его положение в пространстве. Положение объекта затем должно быть сопоставлено с тем местом, где находится рука, и разработана траектория движения к цели. Наконец, пространственная траектория должна быть превращена в координированное сокращение мышц. Какие из этих процессов происходят в первичной моторной коре? Эксперименты по контролируемому перемещению конечностей показывают, что некоторые нейроны в первичной моторной коре в участке, отвечающем за движение руки, разряжаются с максимальной частотой тогда, когда движение совершается в определенном направлении 106). Преимущественное направление не является абсолютным, изменение угла движения приводит не к устранению, а к уменьшению частоты разрядов нейронов. Более того, преимущественное направление зависит от начальной позиции конечности или позиции тела107· 108).
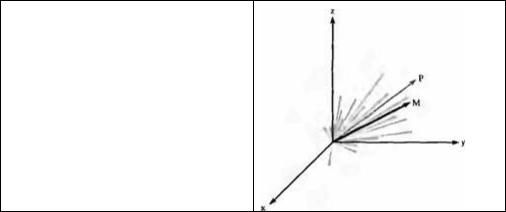
Нейроны зоны M1, отвечающие за сходное направление, образуют кластеры, функционально объединенные возбуждающими синапсами. Нейроны, контролирующие противоположное направление движений, обладают способностью ингибировать друг друга 109). Георгопулус с коллегами 110)--112) предположили, что траектория движения определяется активностью ансамбля нейронов. Выходной сигнал нейронального ансамбля кодирует направление, которое эквивалентно суммарному вектору преимущественного направления отдельных активных нейронов (рис. 22.21). Такое векторное кодирование напоминает способ определения направления в зрительном анализаторе 133).
Планирование движения
Как нейроны в зоне Μ1 определяют пространственную локализацию цели? Шен и Александер114) обучили обезьян совершать движения к предмету, находящемуся либо прямо перед ними, либо располагающемуся под углом в 90° С. При этом им удавалось различать нейрональную активность, которая была либо зависима от положения цели, либо от самого движения. Некоторые нейроны в М1 зоне активировались только во время движения конечностей. Однако многие нейроны активировались сначала при определении локализации цели, а затем при завершении движения. Этот факт предполагает, что трансформация определения пространственной локализации в мышечную активность происходит
Глава 22. Клеточные механизмы двигательного контроля |
509 |
Рис. 22.21. Кодирование движений в моторной коре. Предпочтительное направление движения для кортикальных нейронов, представленное в трех проекциях. Активность каждого нейрона во время отработанных движений конечностей показана как скаляр, длина которого пропорциональна частоте разрядов данного нейрона. Суммарный вектор для группы нейронов (Р) представлен стрелкой, направление которой совладает с направлением движения (М).
Fig. 22.21. Encoding Movement in the Motor Cortex. The preferred direction of cortical neurons is shown in three dimensions. The activity of each neuron during a trained limb movement is plotted as a scalar whose length is proportional to its rate of firing. The vector sum for that population of neurons (P) is an arrow whose direction is similar to that of the movement (M). (After Georgopoulos, Schwartz, and Kettner, 1986.)
в М1 зоне. Маунткасл и коллеги115· 116) использовали специальную тренировку, чтобы научить обезьяну различать частоту тактильных стимулов до того, как совершить определенное движение. Они нашли в Μ1 зоне нейроны, активность которых соответствовала разным характеристикам раздражителя, независимо от последующего движения, что является, повидимому, нейрональным коррелятом аппарата, делающего выбор. Таким образом, Μ1 зона не только прямо управляет мышечным движением, но принимает также участие в нейрональном анализе, который определяет, какой тип движения будет осуществлен.
Понятно, что для передвижения конечности в определенную точку пространства двигательная система должна следовать координатам на нейрональной карте пространства. Такие пространственные карты не могут находиться только в моторной или премоторной зоне. Ассоциативные зоны париетальной коры объединяют вместе сенсорный поток разных модальностей от зрительного, слухового, тактильного и других анализаторов117, 118). Повреждения париетальной коры приводят к синдрому «игнорирования», когда определенные части тела или определенная зона пространства не замечаются119). Этот специфический дефект предполагает, что пространство представлено в различных формах в анатомически различающихся частях мозга.
Двигательный контроль представлен в мозге достаточно широко, включая зону M1, премоторную кору, дополнительную моторную зону и даже соматосенсорную кору121). Так же как для сенсорной системы, эта множественность базируется на наличии параллельных проводящих путей двигательной системы. Это отражает также тот факт, что трудно провести четкую границу между планированием и координированием двигательного акта с одной стороны, и с сенсорным притоком, который необходим для запуска и модуляции движений, — с другой. Действительно, расчет двигательного акта является настолько сложным, что у высших позвоночных эволюционно возникла даже необходимость второго «вычислительного центра» в виде специального образования — мозжечка, предназначенного для сопряжения выполнения двигательной команды с механизмом обратной связи.
§6. Мозжечок
Мозжечок является выростом моста и состоит из трех слоев коры, покрывающих глубокие ядра мозжечка. Его анатомические свойства суммированы в приложении С. Сопоставление структуры мозжечка позвоночных наводит на мысль, что эволюционно мозжечок появился в результате развития вестибулярных ядер122). Действительно, непосредственные соединения мозжечка с первичными вестибулярными афферентами и другими вестибулярными ядрами имеются у всех видов. Однако роль мозжечка в функциях мозга простирается дальше участия в регуляции равновесия и контроля положения тела. Мозжечок также эффективно влияет на неокортекс, он вовлечен в различные формы пластичности и некоторые формы когнитивных процессов124). Однако лучше всего изучена его роль в координации движений. Мозжечковые повреждения характеризуются нарушением координации с незначительными изме-

510 |
|
|
|
|
|
|
|
|
Раздел III. Интегративные механизмы |
|
Рис. |
22.22. Эфферентные |
|
|
|
||||||
и |
афферентные |
пути |
|
|
|
|||||
мозжечка. |
|
|
Полушария |
|
|
|
||||
мозга, |
ствол |
|
мозга |
и |
|
|
|
|||
спинной мозг, показанные |
|
|
|
|||||||
совместно с изображением |
|
|
|
|||||||
верхней |
|
поверхности |
|
|
|
|||||
мозжечка |
|
(справа) |
и |
|
|
|
||||
нижележащих |
|
ядер |
|
|
|
|||||
мозжечка |
|
|
(слева). |
|
|
|
||||
Выходы |
из |
мозжечка |
|
|
|
|||||
(слева, показаны серым) |
|
|
|
|||||||
исходят |
из |
|
зубчатого, |
|
|
|
||||
промежуточного |
и |
ядра |
|
|
|
|||||
шатра. |
|
Волокна |
|
от |
|
|
|
|||
зубчатого |
ядра |
снабжают |
|
|
|
|||||
контралатеральную |
|
|
|
|
|
|||||
моторную |
|
кору |
через |
|
|
|
||||
вентролатеральное |
|
и |
|
|
|
|||||
заднее |
|
|
|
|
|
|
|
|
|
|
вентролатеральное |
ядра |
|
|
|
||||||
таламуса. |
Промежуточное |
|
|
|
||||||
ядро |
|
дает |
проекции |
в |
|
|
|
|||
контралате |
|
ральное |
|
|
|
|||||
красное ядро. Оба этих |
|
|
|
|||||||
тракта |
связаны, |
таким |
|
|
|
|||||
образом, |
с |
латеральной |
|
|
|
|||||
двигательной |
|
системой. |
|
|
|
|||||
|
|
|
||||||||
Ядро |
шатра |
посылает |
|
части мозжечка получают многочисленные входы от спиномозжечкового тракта. |
|
|||||
отростки к вестибулярным |
|
|
|
|||||||
Флоккуло-нодулярная доля снабжается вестибулярными ядрами. |
||||||||||
ядрам |
и |
|
ретикулярной |
|
|
|
||||
формации, |
участвуя |
в |
|
Fig. 22.22. Efferent and Afferent Pathways in the Cerebellum. Cerebral hemispheres, |
||||||
контроле |
|
медиальной |
|
brainstem, and spinal cord, together with a view of the superior surface of the |
||||||
двигательной |
|
системы. |
|
cerebellum (right) and the underlying cerebellar nuclei (left). Outputs from the |
||||||
Входы (справа, показаны |
|
cerebellum (left side, grey) are through the dentate, interposed, and fastigial nuclei. |
||||||||
черным) |
в |
латеральные |
|
Fibers from the dentate nucleus supply the contralateral motor cortex through the |
||||||
полушария |
|
мозжечка |
|
ventrolateral and parts of the ventroposterolateral nuclei of the thalamus. The |
||||||
направляются |
из |
разных |
|
interposed nuclei project to the contralateral red nucleus. Both these pathways, |
||||||
областей |
коры больших |
|
therefore, are associated with the lateral motor system. The fastigial nucleus projects to |
|||||||
полушарий |
мозга |
через |
|
the vestibular nucleus and the pontine and medullary reticular formations, contributing |
||||||
ядра моста. Афферентный |
|
to the medial motor system. Inputs (right side, black) to the lateral hemispheres of the |
||||||||
поток |
от |
красного ядра |
|
cerebellum are from wide areas of the cerebral cortex, through the pontine nuclei. |
||||||
переключается |
в |
нижних |
|
Afferent input from the red nucleus is relayed through the inferior olive. More medially, |
||||||
оливах. |
|
|
|
Более |
|
the cerebellum receives extensive input from the spinocerebellar tracts. The |
||||
медиальные |
|
|
|
|
|
flocculonodular lobe is supplied by the vestibular nucleus. |
||||
нениями силы сокращений мышц или чувствительности. Влияние мозжечка на двигательный контроль опосредовано его мощными связями с премоторной корой, а также, посредством ядер ствола мозга, с двигательными системами спинного мозга.
Мозжечковые связи
Выход из коры мозжечка формируется аксонами клеток Пуркинье, которые образуют тормозные синапсы на нервных клетках глубоких ядер мозжечка или на нейронах вестибулярных ядер. Проекции организованы упорядоченным образом (рис. 22.22): аксоны от флоккулярной и нодулярной доли (вестибулярный мозжечок) направляются непосредственно в вестибулярные ядра125· 126). Остальные пути проецируются в глубокие ядра в медиолатеральной последовательности. Клетки Пуркинье дают проекции в ядро шатра, в промежуточное ядро и, наконец, в зубчатое ядро. Детали связей ядер мозжечка показаны на рис. 22.22. Важной особенностью является то, что зубчатое и промежуточное ядра посылают свои выходы к двигательной коре через вентролатеральное ядро таламуса. Таким образом реализуется важное влияние этих ядер мозжечка на латеральную моторную систему. Промежуточное ядро проецируется также в красное ядро. С другой стороны, ядро шатра проецируется в вестибулярные ядра и ретикулярную формацию, влияя, таким образом, на вестибулоспинальный и ретикулоспинальный тракты, то есть на медиальную

Глава 22. Клеточные механизмы двигательного контроля |
511 |
|
||||
Рис. 22.23. Синаптическая организация мозжечка. Кора |
|
|
|
|||
мозжечка состоит из гранулярных клеток, клеток |
|
|
|
|||
Пуркинье и молекулярного слоя. Единственные |
|
|
|
|||
покидающие кору мозжечка и направляющиеся в |
|
|
|
|||
глубокие ядра мозжечка волокна представлены |
|
|
|
|||
тормозными аксонами клеток Пуркинье. Аксоны |
|
|
|
|||
глубоких ядер мозжечка формируют выходящие пути |
|
|
|
|||
мозжечка. Входы мшистых волокон возбуждают |
|
|
|
|||
гранулярные клетки, аксоны которых достигают |
|
|
|
|||
молекулярного слоя и формируют сеть параллельных |
|
|
|
|||
волокон. |
Параллельные |
волокна |
образуют |
|
|
|
возбуждающие синапсы на клетках Пуркинье, |
|
|
|
|||
звездчатых клетках, корзинчатых клетках и дендритах |
|
|
|
|||
клеток Гольджи. Лазающие волокна образуют |
|
|
|
|||
возбуждающие синапсы на клетках Пуркинье. Как |
|
|
|
|||
мшистые, так и лазающие волокна посылают |
|
|
|
|||
возбуждающие коллатерали к нейронам глубоких ядер |
|
|
|
|||
мозжечка. |
|
|
|
|
|
|
Fig. 22.23. Synaptic Organization of the Cerebellum The |
|
|
|
|||
cerebellar cortex is subdivided into the granule celt Purkinje |
|
|
|
|||
celt and molecular layers. The sole output from the cortical |
fibers form excitatory synapses on |
Purkinje cells, stellate |
||||
layer to cells of the deep cerebellar nuclei is by inhibitory |
||||||
axons from Purkinje cells. The axons of deep nuclear |
cells, basket cells, and dendntes of Golgi cells. Climbing |
|||||
neurons form the output paths of the cerebellum. Mossy- |
fibers form excitatory synapses on Purkinje cells. Both |
|||||
fiber inputs excite granule cells, whose axons ascend to the |
climbing fibers and mossy fibers make excitatory |
|||||
molecular layer to form a parallel fiber network. Parallel |
connections with cells in the deep cerebellar nuclei. |
|||||
моторную систему. Соматотопическое устройство, характерное для коры, продолжается и в нисходящих проекциях.
Входы в мозжечок (рис. 22.22) также представлены раздельно. Латеральные полушария коры мозжечка получают входы от обширных областей коры головного мозга (через ядра моста) и от красного ядра (через нижнюю оливу). Флоккуло-нодулярная (клочково-узелковая) доля получает входы от вестибулярного ядра. Медиальная зона мозжечковой коры получает входы, идущие от проприоцепторов и кожных рецепторов на всех уровнях спинного мозга.
По этой причине Шеррингтон говорил о мозжечке как «головном ганглии проприоцептивной системы». Сенсорные входы образуют соматотопическое представительство в мозжечковой коре, совпадающее с моторным представительством тех же участков тела.
Таким образом, мозжечок получает проприоцептивные, вестибулярные и другие чувствительные входы, а также массивные проекции от моторной и ассоциативной коры, сигнализирующие о выполнении или намерении выполнить произвольное движение.
Клеточное строение коры мозжечка
Кора мозжечка состоит из трех слоев (рис. 22.23) 127, 128). Самый внутренний слой содержит плотно упакованные (общим числом до 1010-1011) гранулярные клетки. Эта величина может соответствовать приблизительной сумме всех других клеток нервной системы! Гранулярные клетки посылают аксоны в наружный молекулярный слой, образуя систему параллельных волокон, каждое из которых тянется до нескольких миллиметров. В слое гранулярных клеток присутствуют также клетки Гольджи, формирующие тормозные синапсы на гранулярных клетках.
Во втором корковом слое находятся клетки Пуркинье, аксоны которых составляют единственный выход из коры мозжечка. Дендриты клеток Пуркинье тянутся в молекулярный слой коры, в сторону параллельных волокон. Параллельные волокна формируют возбуждающие синаптические контакты на шипиках дистальных дендритов клеток Пуркинье. Можно представить себе клетки Пуркинье выстроенными в ряд, как телеграфные столбы, вдоль которых, как проволока, тянутся параллельные волокна. Подсчитано, что каждая клетка Пуркинье получает входы от более чем 200 000 параллельных волокон.
Каждое параллельное волокно объединяет ряд клеток Пуркинье, проецирующихся в определенном порядке к нижележащим ядрам мозжечка. Значение такого объединения состоит в том, что оно может контролировать сразу несколько суставов, например плечо.

512 |
Раздел III. Интегративные механизмы |
Рис. 22.24.
Внутриклеточная регистрация простых и комплексных потенциалов действия
вклетках Пуркинье мозжечка кошки. Комплексный потенциал действия (отмечен звездочкой), возникающий
вответ на стимуляцию лазающих волокон, имеет выраженную фазу «плато»
инаблюдается при
частотах |
значительно |
Fig. 22.24. Intracellular Recording of Simple and Complex Spikes in a cerebella |
меньших, чем те, которые |
Purkinje cell from an anesthetized cat. Complex spikes (star) due to climbing-fiber input |
|
вызываются |
|
have pronounced "plateau potentials" and occur at much lower frequencies than do |
синаптическим |
входом, |
simple spikes caused by mossy fiber-granule cell-parallel fiber input. (After Martinez, |
приходящим от мшистых |
Crilt and Kennedy, 1971.) |
|
волокон |
через |
|
гранулярные |
клетки и |
|
параллельные волокна. |
|
|
локоть и запястье, обеспечивая, таким образом, механизм координации движений в нескольких суставах128). Длина параллельных волокон достаточна для того, чтобы объединить клетки Пуркинье, проецирующиеся на соседние глубокие ядра, что может обеспечить межъядерную координацию. Второй корковый слой имеет также в своем составе звездчатые и корзинчатые клетки, которые обеспечивают тормозные входы к клеткам Пуркинье от отдаленных параллельных волокон. Этот механизм аналогичен латеральному торможению в сенсорных системах.
Входные сигналы в мозжечок от коры и от сенсорных систем поступают по мшистым волокнам, которые создают возбуждающие синапсы с гранулярными клетками, клетками Гольджи и нейронами глубоких ядер. Возбуждение мшистыми волокнами параллельных волокон (аксонов гранулярных клеток) вызывает генерацию простых спайков в клетках Пуркинье (рис. 22.24)129). Это тоническое возбуждение происходит со скоростью от 50 до 150 в секунду. Одиночное лиановидное (лазающее) волокно, идущее от нижней оливы, формирует возбуждающие синапсы на соме и проксимальных дендритах 1-10 клеток Пуркинье. Лиановидные волокна вызывают сильное возбуждение клеток Пуркинье 130, 131), проявляющееся в виде комплексных спайков (рис. 22.24). Эта активность вызывает модулируемые потенциал-активируемыми калиевыми каналами133) кальциевые потенциалы действия в дендритах, что приводит к значительному входу ионов кальция132). Во время генерации комплексных спайков простые спайки обычно подавлены134· 135).
Клеточная активность в ядрах мозжечка
Тач и его коллеги136· 137) изучали взаимоотношения между заученными движениями и клеточной активностью в глубоких ядрах мозжечка. Обезьян учили выполнять ряд последовательных сгибательных и разгибателъных движений с нагрузкой. Изменяющийся груз служил для того, чтобы отделить мышечную активность, связанную с положением сустава, от активности, зависящей от направления движения. Клетки в ядрах мозжечка разряжались в трех вариантах: одни были активны при выполнении работы с нагрузкой, другие были активны при определенном положении сустава, третьи генерировали разряды при движении в определенном направлении.
Во время заученных движений клетки зубчатого ядра активировались раньше, чем клетки первичной моторной коры, затем после моторной коры следовала активность в промежуточном ядре мозжечка, а завершало процесс мышечное сокращение. Эта последовательность согласуется с идеей о том, что информация о планируемых движениях передается от ассоциативной моторной коры к латеральной доле мозжечка, где она обрабатывается и посылается обратно к первичной моторной коре через зубчатое ядро. Сигналы от моторной коры затем передаются к соответствующим спинальным мотонейронам, а также проходят через промежуточное ядро, где возможна поправка двигательной команды.
Идея о том, что зубчатое ядро играет роль в инициации планируемых действий, поддер-