

Nikolls_-_Ot_neyrona_k_mozgu
.pdfГлава 17. Трансдукция механических и химических стимулов |
373 |
Рис. 17.10. Концевые связи в стереоцилии волосковой клетки. (А) Изображение связей между стереоцилиями, полученное с помощью сканирующего электронного микроскопа. (В) Концевые связи расположены так, что отклонение волоскового пучка в возбуждающем направлении растягивает концевую связь и открывает трансдукционный канал (справа), тогда как противоположно направленное движение сжимает концевую связь (слева).
Fig. 17.10. Tip Links on Hair Cell Stereocilia. (A) Scanning electron micrograph showing extracellular fibers that run from the tips of shorter Stereocilia to the sides of adjacent taller Stereocilia. In some cases these appear to bifurcate. (B) Tip' links are positioned so that deflection of the hair bundle in the excitatory direction stretches the tip link and pulls open the transduction channel (right), while the opposite motion compresses the tip link (left). The orientation of the tip link is appropriate to serve as a gating spring that pulls directly on the gate of the mechanical transduction channel. (Micrograph kindly provided by D. Furness.)
Каналы трансдукции в волосковых клетках
Какого типа каналы открываются на концах волосковых клеток? Оказалось, что это неизбирательные катионные каналы, имеющие значительную кальциевую проводимость. Проводимость одиночного канала — около 100 пСм 53· 55· 56). На основании этих данных, а также измерений величины общего генераторного тока можно рассчитать, что каждая волосковая клетка имеет около 100 каналов трансдукции. Это соответствует всего нескольким каналам на один стереоцилий!
Очень малое количество каналов в каждой волосковой клетке существенно затрудняет их биохимическое и молекулярно-биологическое исследование. Каналы трансдукции волосковых клеток не являются потенциалзависимыми и не открываются какими-либо лигандами в традиционном смысле этого слова. Поэтому маловероятно, что возможна жесткая гомологичность между ними и другими классами управляемых ионных каналов, для которых гены уже описаны. Однако все больше механочувствительных ионных каналов клонируется из бактерий, дрожжей, нематод и мух27· 28), так что гены-претенденты, скорее всего, будут определены в ближайшем будущем.
Адаптация волосковых клеток
Волосковые клетки чрезвычайно чувствительны, пороговые ответы в них возникают при смещении волоскового пучка менее чем на 10--9 м. Поэтому кажется весьма вероятным, что некоторые виды адаптивных процессов должны восстанавливать чувствительность в присутствии «фоновых» стимулов. Например, вестибулярные волосковые клетки в маточке и мешочке должны оставаться чувствительными к самым незначительным движениям головы, испытывая постоянное влияние силы тяжести, действующей на покровную отолитовую мембрану. Путем прямого измерения можно показать, что в процессе длительного смещения волосковые клетки действительно адаптируются.
Рис. 17.11 показывает результат эксперимента, в котором постоянное отклонение на 0,65 мкм было приложено к пучку ресничек волосковой клетки черепахи, мембранный потенциал в которой устанавливался на разных уровнях путем фиксации напряжения. При -72 м В входящий трансдукционныи ток сначала достигал на пике 100 пА, а затем спадал с постоянной времени примерно 10 мс. Это адаптивное изменение вызвано притоком ионов кальция через открытые трансдукционные каналы. Когда потенциал мембраны был фиксирован на +60 мВ, то возникал выходящий трансдукционный ток (равновесный потенциал равен 0 мВ), который сохранялся

374 |
Раздел III. Интегративные механизмы |
|
|
Рис. 17.11. Адаптация генераторного тока в волосковых клетках. (А) Схема опыта. (В) Мембранные потенциалы волосковых клеток в процессе механотрансдукции. При отрицательных мембранных потенциалах ток был входящим и спадал во время раздражения. При положительных мембранных потенциалах ток менял направление и не спадал; вход кальция также снижался, устраняя кальций-зависимую адаптацию.
Fig. 17.11. Adaptation of Transducer Current in Hair Cells. (A) A sustained deflection of the hair bundle was imposed with a piezoelectric manipulator, and transducer current was recorded by voltage clamp. (B) Membrane currents recorded from a hair cell during mechanotransduction. At negative membrane potentials the current was inward and decayed during the displacement. At positive membrane potentials the current was reversed in polarity and showed no decay; calcium influx is also reduced, so calcium-dependent adaptation is eliminated. (After Crawford, Evans, and Fettiplace, 1989.)
неизменным, адаптация при этом устранялась. При +60 мВ движущая сила для кальция мала, и в клетку проникает небольшое количество кальция. В других экспериментах адаптация также устранялась удалением кальция из внеклеточного раствора или внутриклеточной инъекцией сильных кальциевых хелаторов57).
В некоторых волосковых клетках адаптация сопровождается изменениями жесткости пучка, что предполагает «переустановку» жесткости воротной пружины58). Это наблюдение, в сочетании с кальциевой зависимостью адаптации, привело к предположению, что движение миозина вдоль актинового каркаса стереоцилии может смещать положение места прикрепления концевой связи, изменяя жесткость пучка59). Путем клонирования миозинов из внутреннего уха60) и использования специфических антител, было показано, что миозин Ιβ (немышечная форма) расположен вблизи концов стереоцилии в волосковых клетках лягушки61). Эта относительно медленная адаптация, основанная на действии миозина, вероятно, является наиболее значимой в вестибулярных волосковых клетках. Более быстрая форма адаптации в слуховых волосковых клетках обусловлена ионами кальция, действующими непосредственно на трансдукционные каналы, заставляя их закрываться62).
Тесная связь между механическим входом и открыванием трансдукционного канала имеет важные последствия для функции внутреннего уха. Такая тесная связь предполагает, что в процессе трансдукции будет возникать отрицательная обратная связь. Таким образом, рецепторный потенциал изменяет вход кальция (через изменение движущей силы), приводя к изменениям жесткости или положения волоскового пучка. Действительно, изменение
мембранного потенциала волосковых клеток черепахи приводит волосковый пучок в движение41).
Глава 17. Трансдукция механических и химических стимулов |
375 |
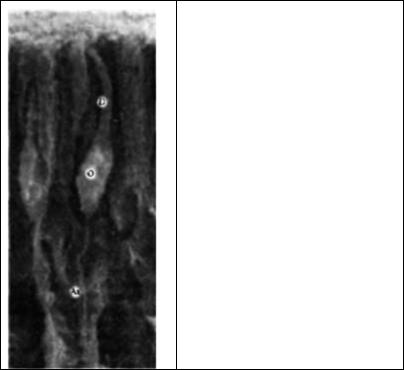
Рис. 17.12. Обонятельные рецепторные нейроны. Электронная микрофотография обонятельного эпителия хомяка.
Fig. 17.12. Olfactory Receptor Neurons. Scanning electron micrograph of the olfactory epithelium of a hamster. Each receptor neuron (0) has a long dendrite (0) that extends to the surface and an axon (Ax) that projects from the epithelium to the olfactory bulb. The long sensory cilia at the tip of the dendrite form a dense mat in this preparation and are not individually resolved. (Micrograph kindly provided by R. Costanzo.)
Активная механика волосковых клеток может способствовать проявлению некоторых необычных свойств слуха, включая звуки (называемые отоакустическими излучениями), которые могут исходить из ушей большинства видов, включая человека63). Способность уха излучать звук дала аудиологам возможность напрямую оценивать функции волосковых клеток,
даже у младенцев или коматозных больных64).
§3. Обоняние
Механотрансдукция в ухе достигает высокой чувствительности благодаря тесной связи энергии стимула и мембранного потенциала волосковой клетки. В противоположность этому, большая чувствительность обоняния, зрения и некоторых форм вкусовых ощущений достигается благодаря химическому усилению — то есть механизму вторичных посредников, в котором ферментативные каскады производят большое количество промежуточных продуктов, тысячекратно увеличивая тем самым эффект единственной активированной рецепторной молекулы.
Обоняние слабо развито у человека по сравнению с собаками, свиньями и бабочками. Но в то же время значительными усилиями (и рекламным временем) создается то, что можно рассматривать как обонятельное поведение человека (если принять во внимание количество сортов мыла, деодорантов, духов, которые призваны обеспечить социально приемлемый личный букет запахов). Обонятельные сигналы существенно важны для человеческого выживания, стимуляции питания, воспроизведения и связи «мать — дитя» 65· 66). Кроме того, обонятельные стимулы имеют сильную способность пробуждать эмоции и извлекать давно хранимые воспоминания (возможно, как наследие эволюции, от использования обонятельных меток для нахождения дома, как у других видов) 67). Навязчивый характер обонятельной памяти был описан Марселем Прустом в «Воспоминании о прошлом»:
Когда от отдаленных воспоминаний ничего не остается; Когда уже люди мертвы;
Когда вещи разбиты и потеряны; Все же остаются еще, более хрупкие. Но гораздо более живые.
Более невещественные.
Но более постоянные, более верные. Запах и вкус остаются в воздухе навсегда, Как призраки, готовые напомнить нам...
Выявление и различение уникальной смеси индивидуальных запахов, связанной с этими воспоминаниями, начинается с семейства молекулярных рецепторов в обонятельных рецепторных нейронах.
Обонятельные рецепторы
У позвоночных животных запахи выявляются структурой, содержащей примерно 100000 обонятельных рецепторных нейронов, аксоны которых проецируются через тонкий участок фронтального черепа (ситовидную пластинку) в обонятельную луковицу (рис. 17.12).
376 |
Раздел III. Интегративные механизмы |
Длинные реснички обонятельных рецепторов достигают носовой полости, где они лежат в слизистом слое (mucus), толщиной у человека около 50 мкм, который полностью замещается каждые 10 минут. Слизистый слой защищает сенсорный эпителий, вымывая потенциально токсичные летучие соединения, и все одоранты должны просачиваться через него, чтобы достигнуть сенсорных ресничек. Одоранто-связывающий белок помогает концентрировать гидрофобные одоранты в этом насыщенном водой слое68). Обонятельные рецепторы необычны тем, что они постоянно сменяются на протяжении жизни животного. Каждый рецептор живет
один или два месяца, и новые рецепторы переходят из слоя базальных клеток в обонятельный эпителий 69).
Обонятельный ответ
Первые отведения обонятельных ответов были сделаны Эдрианом70) и Оттосоном71). Позже были получены доказательства того, что молекулы одоранта взаимодействуют с рецепторами в мембране реснички, вызывая увеличение проводимости, приводящее к деполяризации. Затем потенциалы действия передаются по аксону обонятельной рецепторной клетки в центральную нервную систему. Методом пэтч-клампа были зарегистрированы одорант--индуцированные токи изолированных обонятельных клеток72) и изучены точный временной ход и место возникновения обонятельного ответа.
Пример такого эксперимента на клетке, изолированной из обонятельной слизистой оболочки саламандры73), показан на рис. 17.13. Потенциал мембраны клетки фиксирован на —65 мВ, и раствор, содержащий смесь молекул одоранта (примерно 0,1 ммоль) в 100 ммоль KCl, подается из второй пипетки короткими (35 мс) импульсами давления — сначала на сому, потом на дистальную часть дендритов и ресничек. Раствор, подаваемый из пипетки на сому, вызывает быстрый входящий ток, благодаря локальному увеличению концентрации калия. Временной ход калиевого ответа обусловлен соотношением скоростей аппликации и последующего разведения раствора за счет диффузии внутри ванночки. Второй, меньший и более медленный входящий ток появляется, когда одорант достигает апикального дендрита.
Раствор, подаваемый на апикальный дендрит и реснички, производит лишь незначительный калиевый ответ, поскольку, предположительно, в этой части клетки меньше калиевых каналов. Однако сам одорант вызывает большой входящий ток, который превышает время аппликации на несколько секунд. Эксперимент ясно показывает, что область чувствительности к одоранту
— это дистальный дендрит и реснички, и пролонгированный временной ход дендритного ответа согласуется с идеей, что изменение проводимости обусловлено системой вторичных посредников.
Каналы обонятельных рецепторов, управляемые циклическими нуклеотидами
Деполяризация, вызываемая молекулами пахучих веществ, обусловлена открыванием неселективных катионных каналов74) и дополнительным протеканием тока через кальцийактивируемые хлорные каналы75· 76). Неселективные катионные каналы в обонятельных рецепторах открываются внутриклеточным цАМФ (рис. 17.14), и эти каналы тесно связаны с катионными каналами, открываемыми цГМФ (циклическим гуанозин-монофосфатом) в палочковых фоторецепторах77). Как и фоторецепторный канал папочек (глава 19), обонятельный рецепторный канал не способен ни активироваться, ни инактивироваться изменениями потенциала мембраны. Эти каналы проницаемы для ионов Na+, K+ и Ca2+. Вход кальция может способствовать процессам адаптации. Благодаря очень высокому входному сопротивлению обонятельных рецепторов, для возникновения потенциала действия в них достаточно всего нескольких открытых катионных каналов. Это позволяет предположить, что даже одна молекула одоранта может быть распознана рецептором 78).
Сопряжение рецептора с ионными каналами
Как связывание одоранта с рецептором сопрягается с открыванием цАМФ-зависимых катионных каналов? Существует предположение, что одорант предпочтительно связывается с теми молекулярными рецепторами, которые сопряжены с G-протеином. Активированный G- протеин высвобождает α-субъединицу, которая стимулирует аденилил-циклазу, синтезирующую цАМФ (рис. 17.14). Идентифицировано обширное семейство претендентов на роль генов обонятельного рецептора79),
Глава 17. Трансдукция механических и химических стимулов |
377 |
Рис. 17.13. Ответы изолированных обонятельных клеток саламандры. Пэтч--кламп отведение от целой клетки. (А) При аппликации раствора на тело клетки возникает кратковременный входящий ток, обусловленный повышением концентрации калия. Второй, более медленный и меньший компонент обусловлен действием одоранта, достигающего удаленных дендритов в процессе диффузии. (В) При аппликации раствора на дендрит первый компонент исчезающе мал, а второй медленный компонент возрастает и длится в течение нескольких секунд после отмывки раствора одоранта.
Fig. 17.13. Responses of Isolated Olfactory Cells from the salamander. A patch clamp electrode is used to record whole-cell current. A solution containing 0,1 mW odorant mixture in 100 mM KCl is applied to the cell by a brief (35 ms) pressure pulse. (A) When solution is applied to the cell body, there is a rapid transient inward current due to the increased KCl concentration, followed by a smaller, slower current as the odorant reaches the apical end of the dendrite. The time course of the fast inward current is indicative of the time course of application and dissipation of the electrode solution. (B) When the solution is applied to the dendrite, there is only a small rapid current due to the KCL but a large current due to the odorant lasts for several seconds after the electrode solution has washed away. (After Firestein, Shepherd, and Werblin, 1990.)
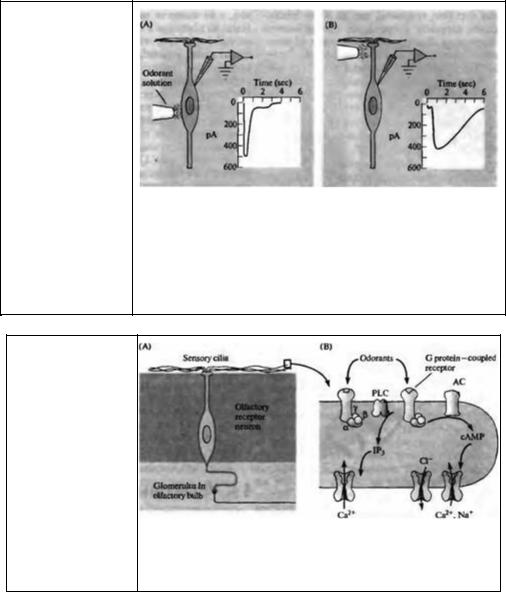
Рис. 17.14. Трансдукция в обонятельной ресничке. (А) Схема восприятия запаха. (В) Система вторичных посредников в обонятельной ресничке.
Fig. 17.14. |
Transduction in |
||||
Olfactory |
Cilia. |
(A) |
The |
||
molecular |
|
receptors |
for |
||
olfactants are found in sensory |
|||||
cilia that project into the |
|||||
mucous layer of the olfactory |
|||||
epithelium. |
|
Depolarizing |
|||
receptor |
potentials |
in these |
|||
long receptors give rise to |
|||||
action |
potentials |
|
that |
||
propagate |
along |
the |
olfactory in the plasma membrane of the olfactory cilia. This frees the a subunit to activate |
||
receptor neuron's axon into the adenylyl cyclase (AC) and raise the concentration of cAMP, which causes nonselective |
|||||
central nervous |
system. |
(B) cation channels to open, depolarizing the membrane. Calcium-gated chloride current |
|||
Odorant |
molecules |
bind |
to can enhance this effect. Other pathways may involve the activation of phospholipase С |
||
specific |
G |
protein-coupled (PLC) and the consequent rise in IP3 to act directly on plasma membrane calcium |
|||
receptors |
|
|
|
|
channels. |
|
|
|
|
|
|
которые кодируют трансмембранные белки, сопряженные с G-протеином, сходные по структуре с метаботропными нейрональными рецепторами (глава 10). Величайшее разнообразие кодируемых белков возникает за счет последовательности трех трансмембранных доменов, которые формируют лиганд-связующий карман. Был идентифицирован80) G-протеин, специфично экспрессируемый в обонятельном эпителии (Golf). Высвобождение Golfα при связывании одорантов вызывает повышение активности
обонятельной аденилил-циклазы, которая содержится в большом количестве в обонятельных ресничках81· 82).
Об эффективности этих путей трансдукции свидетельствует быстрое появление цАМФ. Брир и его коллеги 83), используя ме-

378 |
Раздел III. Интегративные механизмы |
тод stop-flow, показали, что за 50 мс аппликации одоранта в препарате изолированной обонятельной реснички происходит 10-кратное увеличение концентрации цАМФ. Существуют также подтверждения того, что обонятельные нейроны используют в процессе трансдукции G- протеиновую активацию фосфолипазы С и продуцируют IР3 (инозитол трифосфат)84). В этом случае IР3 может напрямую открывать кальциевые каналы в плазматической мембране85). IР3 особенно важен для обоняния у беспозвоночных86). Роль 1Р3 в обонянии млекопитающих менее ясна, о чем можно судить на том основании, что у трансгенных мышей, лишенных цАМФ-регулируемых каналов, полностью отсутствует способность различать запахи87).
Специфичность одорантов
Люди могут различать очень большое число запахов, и существование сотен, а возможно, и тысяч обонятельных рецепторных молекул может служить предпосылкой этой способности. Проблема лишь в отсутствии одорантной специфичности индивидуальных обонятельных рецепторньгх нейронов, каждый из которых распознает определенный спектр запахов, а не является высоко избирательным88). Один из подходов к пониманию этого вопроса состоит в том, чтобы методом in situ гибридизации определить паттерн экспрессии рецепторных молекул, клонируемых обонятельными рецепторными нейронами. Рецептор каждого конкретного одоранта локализован в строго ограниченной области обонятельного эпителия89· 90). Различные семейства рецепторных генов оказались экспрессированными в зонах, вытянутых по длине эпителия (рис. 17.15), при этом каждый конкретный ген экспрессировался ограниченным числом обонятельных рецепторных нейронов.
В дополнение к основному обонятельному эпителию, млекопитающие обладают сошниковоносовым (vomeronasal) органом, который участвует в различении феромонов, стимулирующих спаривание и другие виды поведения. Вомероназальные рецепторные нейроны проецируются в дополнительную обонятельную луковицу, нейроны которой, в свою очередь, проецируются в лимбическую систему. Вомероназальные нейроны экспрессируют два дополнительных семейства обонятельных рецепторов. В них имеются также G-протеин-сопряженные рецепторы, но они отличаются от тех, которые экспрессируются в основном обонятельном эпителии. Каждый вомероназальный нейрон может экспрессировать лишь один тип молекулярного рецептора, при этом паттерн экспрессии у самок и самцов крыс различен91).
§ 4. Механизмы вкуса
Описание механизмов вкуса и запаха часто объединяют, потому что оба ощущения активируются химическими стимулами, приходящими из внешнего мира. Действительно, некоторые вкусовые стимулы действуют на G-протеин сопряженные рецепторы способами, весьма схожими с теми, которые описаны в случае обоняния. Однако другие вкусовые стимулы, в основном соли и кислоты, действуют непосредственно на проводимость мембраны. Кроме того, вкусовые рецепторные клетки анатомически отличаются от обонятельных рецепторных нейронов.
Вкусовые рецепторные клетки
Вкусовые рецепторные являются волосковыми нейроэпителиальными клетками,
Рис. 17.15. (Рис. 17.15В см. на цветной вклейке после с. 640.) Экспрессия генов рецепторов специфических одорантов в подгруппах обонятельных рецепторных нейронов. (А) Обонятельный эпителий крысы, расположенный в складках носовой полости. (В) Полоски обонятельных рецепторных нейронов, меченных специфичными антителами.
Fig. 17.15. Expression of Specific Odorant Receptor Genes by Subsets of Olfactory Receptor Neurons. (A) The olfactory epithelium of a rat lies on a series of convolutions of the nasal cavity called turbinates-Ubeled I to IV. (B) Bands of olfactory receptor neurons label positively for olfactory receptor mRNAs. Probes for different mRNAs label nonoverlapping populations of neurons (depicted as green, yellow, blue, and white dots). (After Vassar, Ngai, and AxeL 1993; В kindly provided by R. Vassar.)

Глава 17. Трансдущия механических и химических стимулов |
379 |
Рис 17.16. Рецепторные вкусовые клетки во вкусовом сосочке. (А) Электронная микрофотография вкусового сосочка в языке крысы. (В) Изолированный вкусовой сосочек, в котором рецепторные клетки помечены антителами к гастдуцину.
Fig. 17.16. Taste Receptor Cells Are Found in Taste Buds within the lingual epithelium. (A)
Transmission electron micrograph of a taste bud in rat tongue. Individual taste receptor cells have microvilli that project into the taste pore to sample the saliva. (B) An individual taste bud dissected from rat tongue. The taste receptor cells (grey) are labeled with an antibody to gustducin, a G protein involved in taste transduction. (A kindly provided by R.Yang and J. Kinnamon, В kindly provided by I.
Wanner and S. D. Roper.)
расположенными во вкусовых сосочках на поверхности языка (рис. 17.16). Как и обонятельные рецепторы, вкусовые клетки регенерируют на протяжении жизни. В отличие от обонятельных рецепторов, вкусовые клетки не имеют аксонов, но формируют химические синапсы с афферентными нейронами во вкусовых сосочках. Микроворсинки направляются от апикального полюса вкусовой клетки в открытую пору вкусового сосочка, где они вступают в контакт с вкусовыми стимулами, растворенными в слюне на поверхности языка.
Вкусовые стимулы обычно подразделяются на пять категорий: соленые, кислые, горькие, сладкие и «умами» — последняя категория обозначается японским словом, обозначающим вкус глутамата натрия, или, в более общем виде, вкус аминокислот. Это многообразие молекул вкусовых стимулов, от простых ионов до сложных карбогидратов и протеинов, сопряжено с широким многообразием механизмов трансдукции. Последние подразделяются на две обширные категории: прямое действие на ионные каналы и опосредованная рецепция вкусовых стимулов, вовлекающая систему вторичных посредников, включая G-протеины92).
Соленый и кислый вкус
Общепринято, что соленый вкус передается непосредственным током натрия (или других одновалентных катионов) по каналам в апикальной мембране вкусовой клетки, открытым в состоянии покоя (рис. 17.17А)93). Натрий присутствует в более высокой концентрации в соленой пище (> 100 ммоль), чем в слюне, поэтому он просто диффундирует внутрь вкусовых клеток по своему электрохимическому градиенту. Возникающая в результате этого деполяризация приводит к выбросу нейромедиатора в химических синапсах, образованных вкусовыми клетками на отростках афферентных нейронов. Натриевые каналы вкусовых клеток не являются потенциалзависимыми, они сходны с эпителиальными натриевыми каналами, расположенными в коже и почках лягушки. Эти натриевые каналы блокируются диуретиком амилоридом (amiloride), и состоят из трех гомологичных субъединиц94). Субъединица а была обнаружена в эпителии языка95).
Кислый вкус вызывается высокой концентрацией протонов в кислых продуктах, которые могут проникать во вкусовые клетки через амилорид-блокируемые каналы (рис. 17.17В)96). Другой механизм, ведущий к деполяризации, является следствием блокады K+ каналов протонами97). Вкусовые клетки лягушки имеют катионные каналы, которые открываются протонами98) и также вызывают деполяризацию. В дополнение к воздействию на реснички вкусовых клеток, соли и протоны могут проникать через вкусовые

380 |
Раздел III. Интегративные механизмы |
Рис. 17.17. Механизм вкусовой трансдукции. Ионный механизм действия рецепторов, чувствительных к (А) соленому, (В) кислому, (С) горькому и (D) сладкому (а также «умами» и горькому).
Fig. 17.17. Mechanisms of Taste
Transduction. Testant molecules range from protons (acids) to simple salts to complex organic compounds. This wide range of chemical stimuli is transduced by a multiplicity of mechanisms. Salts (A) and acids (B) can permeate ion channels in the sensitive ending or block normally open potassium channels.
(C) Some bitter compounds block potassium channels to cause depolarization. (D) Sugars and amino acids (umami) interact with G protein-coupled receptors to initiate second messenger cascades. All these mechanisms lead eventually to depolarizatioa voltagegated calcium influx, and increased release of transmitter onto associated afferent dendrites.
поры (околоклеточный путь) к базолатеральным участкам мембраны рецепторной клетки, действуя там на те же или другие ионные каналы (включая и такие, которые являются амилорид-нечувствительными)99). Этот механизм, вероятно, является общим принципом вкусового восприятия: для каждого отдельного класса вкусовых веществ имеется несколько параллельных путей трансдукции.
Сладкий и горький вкус
Сладкие вещества (сахар) и горькие вещества (часто это растительные алкалоиды, прототипом которых является хинин) обычно представляют собой крупные молекулы, которые связываются с макромолекулярными рецепторами с высокой степенью специфичности. За немногими исключениями, сладкие и горькие вещества активируют систему вторичных посредников путем взаимодействия с G-протеин сопряженными рецепторами (рис. 17.17С и D). Только немногие из этих макромолекулярных вкусовых рецепторов были идентифицированы (см. следующий раздел), но G-протеин, специфичный для вкусовой клетки, густдуцин (gustducin), был клонирован и оказался гомологичен трансдуцину — фоторецепторному G-протеину100). Во вкусовых клетках обнаружен широкий спектр молекул, участвующих в последующих каскадах, включая цАМФ фосфодиестеразу, фосфолипазу С и другие компоненты путей, ведущих к синтезу циклических нуклеотидов и IР392). G-протеиновая гипотеза горького и сладкого вкуса подтверждается наблюдением, что трансгенные мыши, лишенные гена густдуцина, могли ощущать соленое и кислое, но не горькое и сладкое 101).
Молекулярные рецепторы для глутамата и чили
Свободный глутамат встречается во многих продуктах, включая мясо, сыр и некоторые овощи, и используется как пищевая приправа в форме глутамата натрия (ГН). Вкус глутамата передается G-протеин-сопряженным метаботропным глутаматным рецептором, который специфически экспрессируется во вкусовых сосочках, но не в окружающем нечувствительном языковом эпителии102· 103).
Методом условной вкусовой аверсии было показано, что как ГН, так и специфичный агонист mGluR4 рецепторов (метаботропных глутаматных рецепторов 4-го типа) L-AP4, вызывают сходные вкусовые ощущения у крыс.
Глава 17. Трансдукция механических и химических стимулов |
381 |
В таких исследованиях используют тесную связь между вкусом продукта и вызванной им тошнотой, чтобы определить, подобны ли вкусовые ощущения. Крысы, у которых после вкусовых ощущений, обусловленных ГН, вызывали тошноту (внутрибрюшинной инъекцией LiCl), затем боялись вкуса mGIuR4-специфичного агониста L-AP4 (а также ГН), но не специфичных агонистов других глутаматных рецепторов.
Таким образом, молекулярный рецептор глутамата является специфическим вкусовым рецептором на языке. «Жгучий» вкус перцев чили представляет другой пример многофункциональности молекулярных рецепторов. Жгучие перцы чили не воспринимаются самими вкусовыми клетками, но скорее болевыми волокнами в языке, которые активируются капсаициновыми соединениями. Рецептор капсаицина клонирован, и доказано, что это кальций-селективный катионный канал104). Рецептор «жгучести» (обозначенный VRI, потому что он был первым из членов рецепторного семейства ваниллоидов) образован волокнами мелкого размера (С-волокна) от клеток спинальных ганглиев, сигнализирующих о боли. Таким образом, природа снабдила перцы чили химической нацеленностью на данный рецептор, возможно, чтобы отпугивать травоядных путем активации болевых волокон, — что оказалось не столь успешной стратегией в случае с человеком, с его пристрастием к острой пище.
§ 5. Трансдукция ноцицептивных и температурных стимулов
При температуре около 33° С у нас обычно не возникает температурных ощущений. Когда температура кожи поднимается или опускается выше или ниже этой нейтральной точки, это вызывает ощущение тепла или холода. Существуют два вида температурных рецепторов в коже: один сигнализирует о тепле, другой — о холоде. Вы можете легко продемонстрировать это на тыльной стороне кисти руки: если касаться кожи в различных местах каким-либо предметом, имеющим комнатную температуру (например, острием обычного карандаша), то в некоторых точках это вызовет ощущение холода. Вне этих точек ощущается только прикосновение. Чтобы обнаружить другие точки, пространственно отличающиеся от Холодовых, где ощущается тепло, используется теплый металлический пробник. Таких мест меньше, и их нужно долго искать. Кожные температурные афференты отличаются от тех, которые сигнализируют об экстремальных температурных воздействиях, вызывающих болевые ощущения.
Активация и сенситизация ноцицепторов
Ноцицепция (восприятие стимулов боли и повреждения) возникает из сочетания прямых и непрямых воздействий на периферические сенсоры. Болезненное нагревание (выше 43° С) вызывает открывание неспецифических катионных каналов в окончаниях С- волокон105· 106). Ионы кальция и натрия входят в клетку и деполяризируют ее, вызывая генерацию потенциала действия. Кислоты могут также напрямую открывать катионные каналы. Кислотно-чувствительные ионные каналы (ASIC) были клонированы из ноцицептивных нейронов107). Механическое повреждение кожи может привести непосредственно к деполяризации ноцицепторов108). Когда клетки, содержащие капсаициновые рецепторы VR1, быстро нагреваются, то открывание этих рецепторов приводит к возникновению болезненного теплового ощущения. Продолжительное воздействие капсаицина вызывает постепенное накопление кальция и смерть клетки. Парадоксально, но капсаицин используется и как долговременный анальгетик, поскольку, убивая С-волоконные афференты, он облегчает тем самым хронические боли 109).
Помимо болевых стимулов, таких как тепло или кислота, которые могут воздействовать непосредственно на ноцицепторы, поврежденные клетки выделяют химические активаторы, такие как аденозинтрифосфат (АТФ). Одна из субъединиц АТФ-рецептора (Р2Х3) специфична для клеток ганглиев задних корешков, образующих С-волокна, и может объединяться с другими субъединицами, вызывая медленно десенситизирующееся возбуждение ноцицепторов аденозинтрифосфатом110)--112). Клеточное повреждение также ведет к выделению цитоплазматических протеаз, которые затем расщепляют белки плазмы крови. Таким образом, из кининогена, распространенного неактивного предшественника, производится пептид
382 |
Раздел III. Интегративные механизмы |
брадикинин, состоящий из 9 аминокислотных остатков. Брадикинин является сильным активатором окончаний С-волокон, вызывающим в них входящий ток и генерацию потенциалов действия. Рецептор брадикинина может действовать через повышение уровня вторичного посредника IР3 113).
Брадикинин и другие химические вещества в поврежденной коже также повышают чувствительность ноцицептивных окончаний. Активируемые теплом катионные токи больше по величине и возникают при более низких температурах, как результат активации брадикинином протеинкиназы С30). Другими медиаторами воспалительного процесса являются простагландины, серотонин, гистамин и субстанция Р. Простагландин Е2 и серотонин повышают уровень цАМФ, увеличивая тем самым амплитуду и потенциалочувствительность натриевого тока в ноцицепторах 114). Повреждение тканей также повышает экспрессию α-адренергических рецепторов в нейронах ганглиев задних корешков — это еще один из механизмов увеличения возбудимости115). Активированные болевые волокна выделяют вещество Ρ не только из своих синапсов внутри спинного мозга (глава 14), но также из своих терминалей в коже. На периферии вещество Ρ может увеличивать возбудимость С-волокон, блокируя К+ каналы116). Процесс сенситизации сопровождается локальной вазодилятацией и опуханием. Поврежденная область становится «гиперальгезивной», т. е. приобретает пониженный болевой порог.
Выводы
·Каждый тип сенсорных рецепторов отвечает предпочтительно на один тип стимула — адекватный стимул.
·Короткие и длинные рецепторы различаются морфологически и функционально. Короткие рецепторы кодируют интенсивность стимула непосредственно в амплитуде рецепторного потенциала. Длинные рецепторы, помимо этого, преобразуют амплитуду рецепторного потенциала в частотный код импульсной активности.
·Ответ многих рецепторов нелинейно зависит от интенсивности стимула. Это обеспечивает некоторым типам рецепторов широкий динамический диапазон.
·Наиболее чувствительные рецепторы адаптируются в той или иной степени во время длительного стимула. Адаптация обусловлена как механическими, так и электрическими факторами. Некоторые рецепторы за счет очень быстрой адаптации «настраиваются» на быстро изменяющиеся стимулы, такие как вибрация.
·Механочувствительные волосковые клетки внутреннего уха непосредственно преобразуют движение в открытие ионных каналов посредством физической связи. Концевая связь, которая соединяет соседние стереоцилии, растягивается посредством отклонения волоскового пучка, открывая таким образом ионный канал.
·Вход кальция через неселективный канал механотрансдукции волосковых клеток приводит к адаптации и закрыванию канала.
·Обонятельные нейроны используют G--протеин-сопряженные мембранные рецепторы, что ведет к открыванию цАМФ--управляемых катионных каналов в плазматической мембране. Каждый представитель обширного семейства обонятельных рецепторных белков экспрессируется небольшим количеством клеток. Все нейроны, экспрессирующие конкретный рецепторный белок, проецируются в единую гломерулу обонятельной луковицы.
·Аминокислоты, сахара и горькие соединения связаны с G-протеин-сопряженными рецепторами во вкусовых сенсорных клетках. Соль и протоны (кислые соединения) воздействуют непосредственно на ионные каналы, генерируя рецепторные
потенциалы |
во |
вкусовых клетках. |
· Болевые |
и |
температурные ощущения передаются разнообразными химическими |
посредниками. Прямое механическое повреждение или чрезмерное нагревание вызывает потенциал действия в болевых волокнах. Соединения, выделяемые из поврежденных тканей, такие как брадикинин, сенситизируют нонцицептивные окончания.