

Передача сигнала рецепторами, связанными с g-белками проходит с участием вторичных посредников
Механизм проведения информационного сигнала с участием рецепторов, связанных с G-белками, состоит, по крайней мере, из 5 этапов.
Первый этаппередачи сигнала включает распознавание и связывание сигнальной молекулы специфическим рецептором плазматической мембраны клетки-мишени.
Второй этаппредставляет собой сложный процесс переноса информации с внешней поверхности плазматической мембраны клетки на внутреннюю, что приводит кG-белок-зависимой активации мембрано-связанных ферментов (например, аденилатциклазы или фосфолипазы С).
Третий этапэтого многоступенчатого каскада сопровождается образованием и повышением уровня в цитозоле одного или нескольких низкомолекулярных внутриклеточных посредников (цАМФ, цГМФ, ДАГ, ионы Са2+ и т.д.).
Повышение уровня низкомолекулярных внутриклеточных посредников внутри клетки инициирует четвертый этаппроведения внешнего регуляторного сигнала – подключение ферментов, активность которых непосредственно регулируется низкомолекулярными внутриклеточными посредниками. Это, как правило, протеинкиназы, которые в свою очередь (пятый этаппроведения регуляторного сигнала) оказывают влияние на активность ключевых клеточных ферментов, определяющих суть ответной реакции клетки на полученный ею извне конкретный информационный сигнал. Активированные каталитические субъединицы некоторых протеинкиназ (например, цАМФ – зависимых) могут оказывать свое влияние непосредственно в цитозоле или транслоцироваться из цитоплазмы в ядро при различных функциональных состояниях клетки. Результатом внутриядерных процессов может быть активация или торможение клеточного цикла, а также синтеза белковых молекул.
Образование вторичных посредников – дело интегральных белков плазматической мембраны.
Первые этапы передачи сигнала с образованием вторичных посредников протекают с участием интегральных белков плазматической мембраны. В этом процессе участвуют 7 ТМС рецептор, Gбелок и фермент, способствующий образованию вторичного посредника.
С хематическое
изображение структуры-адренэргического
рецептора показана на рис слева. Этот
рецептор является членом одного из
подсемейств большого суперсемейства
7 ТМС рецепторов, включающего 3 подсемейства:
1) подсемейство-адренергического
рецептора; 2) подсемейство секретиноподобных
рецеторов;3)подсемейство
метаботропнык глутаматных рецепторов.
Это разделение основано на особенностях
аминокислотных последовательностей
этих рецепторов.
хематическое
изображение структуры-адренэргического
рецептора показана на рис слева. Этот
рецептор является членом одного из
подсемейств большого суперсемейства
7 ТМС рецепторов, включающего 3 подсемейства:
1) подсемейство-адренергического
рецептора; 2) подсемейство секретиноподобных
рецеторов;3)подсемейство
метаботропнык глутаматных рецепторов.
Это разделение основано на особенностях
аминокислотных последовательностей
этих рецепторов.
Представители подсемейства -адренергического рецептора составляют большинство 7-TMS рецепторов, идентифицированных к настоящему времени. Это подсемейство включает- и-адренергические рецепторы, рецепторы гистамина, серотонина, брадикинина, субстанции Р, ангиотензина, интерлейкина 8, тромбина, саматостатина, гликопротеиновых гормонов, ацетилхолина (М-холинорецепторм) и т.д.
-адренергический рецептор, связывающий катехоламиновые агонисты (адреналин, норадреналин и т.д.), состоит из 418 аминокислотных остатков. Его внеклеточный N-концевой домен имеет два участка (сайта) гликозилирования. Гидрофобные участки аминокислотной последовательности -адренергического рецептора образуют 7 трансмембранных а-спиральных сегментов, которые соединены между собой гидрофильными экстраклеточными и внутриклеточными петлями (последние содержат участки взаимодействия с G-белком).
Подсемейство секретиноподобных рецепторов.Это подсемейство включает пептидергические рецепторы секретина, глюкагона, вазоинтестинального пептида, паратирина, кальцитонина, соматолиберина и др. Эти рецепторы характеризуются достаточно высокой гомологией аминокислотной последовательности внутри подсемейства и низкой гомологией с рецепторами подсемейства-адренергического рецептора. Все представители подсемейства секретиноподобных рецепторов связаны с Gs-белком, активация которого лигандорецепторным комплексом приводит к увеличению внутриклеточного уровня цАМФ. Вместе с тем, известно, что наряду с активацией Gs-белка, рецепторы глюкагона, паратирина и кальцитонина активируют и другие G-белки, приводящие к увеличению внутриклеточного уровня Са2* и инозитолтрифосфата.
Подсемейство метаботропных глутаматных рецепторов.Метаботропные глутаматные рецепторы образуют третье, высокоспециализированное подсемейство 7-TMSрецепторов. Глутаматорецепторный комплекс активирует Gq-белок, что в конечном итоге приводит к стимуляции образования инозитолтрифосфата и мобилизации внутриклеточного Са2*. Поскольку увеличение внутриклеточного содержания последнего ведет к активации многих метаболических процессов в клетке эти рецепторы носят название метаботропных в отличие от ионотропных глутаматных рецепторов, являющихся лигандозависимыми ионными каналами. Метаботропные глутаматные рецепторы присутствуют в клетках центральной и периферической нервной системы, где участвуют в регуляции разнообразных функций в ответ на действие агониста L-глутамата.
 Метаботропные
рецепторы характеризуются очень низкой
степенью гомологии аминокислотной
последовательности со всеми другими
7-TMS рецепторами. Эти рецепторы имеют
значительно более высокую молекулярную
массу по сравнению с типичными
представителями других подсемейств
7-ТМSрецепторов.
Метаботропные
рецепторы характеризуются очень низкой
степенью гомологии аминокислотной
последовательности со всеми другими
7-TMS рецепторами. Эти рецепторы имеют
значительно более высокую молекулярную
массу по сравнению с типичными
представителями других подсемейств
7-ТМSрецепторов.
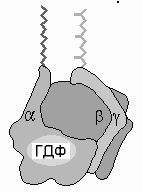
Рис.11.5. Схема строения мембраносвязанного Gбелка
Второй участник передачи сигнала – G белки. Эти белки довольно широко представлены в клетке. Можно выделить по крайней мере 2 типаGбелков:
а) большие, состоящие из нескольких субъединиц, (как правило, гетеротримеры, состоящие из ,исубъединиц с молекулярной массой 42kDa, 35kDaи 10kDaсоответственно) обычно связаны с мембранами благодаря липидным компонентам (пальмитиновая кислота) ковалентно присоединяемым к белку после синтеза на рибосомах
б) низкомолекулярные, состоящие из одной полипептидной цепи (23 кДа), находятся в цитозоле
Все Gбелки обладают ГТФазной активностью (отсюда их название) и их конформация зависит от того, связаны ли они в данный момент с ГДФ или ГТФ. Эти белки в клетке играют роль своеобразных молекулярных «выключателей» многих биохимических процессов (см. раздел по синтезу белков).
 На рис.11.5 представлена схема строения
мембраносвязанногоGбелка. Он состоит из 3 субъединиц,и.
В обычном (неактивном) состоянии три
субъединицы соединены вместе исубъединица связана с ГДФ.
На рис.11.5 представлена схема строения
мембраносвязанногоGбелка. Он состоит из 3 субъединиц,и.
В обычном (неактивном) состоянии три
субъединицы соединены вместе исубъединица связана с ГДФ.
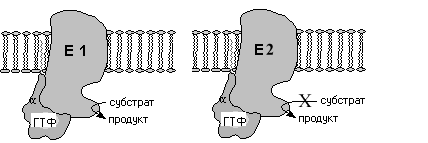 После присоединения гормона к 7 ТМС
рецептору и взаимодействия сGбелком в последнем происходит замена
ГДФ на ГТФ иGбелок
диссоциирует с образованием свободнойсубъединицы и димера субъединиц.субъединица и димерсубъединиц затем перемещаются по
мембране и могут взаимодействовать с
третьим мембраносвязанным участником
передачи сигнала эффекторным белком.
Им может быть фермент, катализирующий
образование низкомолекулярных вторичных
посредников.
После присоединения гормона к 7 ТМС
рецептору и взаимодействия сGбелком в последнем происходит замена
ГДФ на ГТФ иGбелок
диссоциирует с образованием свободнойсубъединицы и димера субъединиц.субъединица и димерсубъединиц затем перемещаются по
мембране и могут взаимодействовать с
третьим мембраносвязанным участником
передачи сигнала эффекторным белком.
Им может быть фермент, катализирующий
образование низкомолекулярных вторичных
посредников.
Конечный эффект такого взаимодействия или активирование (Gs–белок) или ингибирование (Gi-белок) эффекторного белка. Продолжительность эффекта определяется ГТФазной активностью-субъединицы и после гидролиза ГТФ до ГДФ действие субъединиц прекращается и происходит реассоциация их в исходное тримерное состояние.