

77. Углеводы, их биологическая роль, классификация и номенклатура. Основные представители углеводов. Основные пути обмена углеводов. Гликолиз и глюконеогенез
Общая характеристика углеводов. К классу углеводов относят органические соединения, содержащие альдегидную или кетонную группу и несколько спиртовых гидроксилов. Их элементарный состав выражается общей формулой СmН2nOn,. Из этого правила есть немногочисленные исключения, и хотя указанное определение не является абсолютно точным, оно позволяет наиболее просто характеризовать эту группу разнообразных органических соединений в целом. Углеводы служат материалом, при окислении которого выделяется энергия, необходимая для осуществления химических реакций. Такие углеводы рассматривают как резервные. Наряду с этим промежуточные продукты окисления углеводов используются для синтеза многих других органических соединений. Перечисленные функции углеводов (структурная, энергетическая и метаболическая) рассматривают как канонические. Кроме того, многие углеводы и углеводсодержащие биополимеры обладают уникальным строением и специфичностью. Так, групповые вещества крови, являющиеся гликопротеинами, где 80% молекулы представлены углеводами, за счет асимметрических центров, стереоизомеров, таутомеров и конформеров последних приобретают специфичность взаимодействия. Олигосахаридные фрагменты гликопротеинов и гликолипидов клеточных стенок выполняют рецепторные функции. В частности, при их посредстве с клетками связываются белковые токсины (например, холерный, ботулический, и др.), бактерии (кишечная палочка), вирусы (вирус гриппа) и т. п. Структуры олигосахаридных фрагментов иммуноглобулинов высоко воспроизводимы и умеренно консервативны, что обеспечивает специфические углеводно-белковые взаимодействия между доменами этих удивительно тонко организованных защитных белков. Часть ферментов обладают олигосахаридпыми фрагментами. Углеводы делят на две группы: простые и сложные. Простые не подвергаются гидролизу. Сложные углеводы при гидролизе распадаются с образованием простых углеводов.
Простые углеводы. Подавляющее большинство простых углеводов имеет состав СnП2nОn. Ввиду того что простые углеводы не гидролизуются, их называют моносахаридами.
Моносахариды — твердые вещества, легко растворимые в воде, плохо — в спирте и совсем нерастворимые в эфире. Водные растворы имеют нейтральную реакцию на лакмус. Большинство моносахаридов обладает сладким вкусом. В свободном виде в природе встречается преимущественно глюкоза. Она же является структурной единицей многих полисахаридов. Другие моносахариды в свободном состоянии встречаются редко и в основном известны как
компоненты олиго- и полисахаридов. Моносахариды могут существовать как в открытой (оксоформе), так и циклических формах. В растворе эти изомерные формы находятся в динамическом равновесии.
О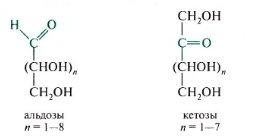 ткрытые
формы:
Моносахариды (монозы) являются
гетерофункциональными соединениями.
В их молекулах одновременно содержатся
карбонильная (альдегидная или кетонная)
и несколько
ткрытые
формы:
Моносахариды (монозы) являются
гетерофункциональными соединениями.
В их молекулах одновременно содержатся
карбонильная (альдегидная или кетонная)
и несколько
гидроксильных фупп, т. е. моносахариды представляют собой полигидроксикарбонильные соединения — полигидроксиалъдегиды и полигидроксикетоны. Для них характерно наличие неразветвленной углеродной цепи.
Моносахариды классифицируют с учетом двух признаков:
• природы карбонильной группы;
• длины углеродной цепи.
Моносахариды, содержащие альдегидную группу, называются альдозами, кетонную группу (обычно в положении 2) — кетозами (суффикс -оза характерен для названий всех моносахаридов: глюкоза, галактоза, фруктоза).
Стереоизомерия. Молекулы моносахаридов содержат несколько центров хиральности, что служит причиной существования большого числа стерсоизомеров, соответствующих одной и той же структурной формуле. Например, в альдогексозе имеются четыре хиральных атома углерода и ей соответствуют 16 стереоизомеров. Кетогексозы содержат по сравнению с соответствующими альдозами на один хиральный атом углерода меньше, поэтому число стерсоизомеров (23) уменьшается до 8 (4 пары энантиомеров). Открытые (незамкнутые) формы моносахаридов изображают в виде проекционных формул Фишера Углеродная цепь в них записывается вертикатьно. У альдоз наверху помещают альдегидную группу, у кетоз — соседнюю с карбонильной первично-спиртовую группу. С этих групп начинают нумерацию цепи. К часто встречающимся алыюпентозам относятся D-рибоза, D-ксилоза, к кетонентозам — D-рибулоза и D-ксилулоза. Наиболее широко распространенными в природе являются альдогсксозы — D-глюкоза, D-галактоза, D-манноза, а кетогексозы часто бывают представлены D-фруктозой.
Относительная конфигурация моносахаридов определяется по конфигурационному стандарту — глицериновому альдегиду. С конфигурацией его хиралыюго центра сравнивается конфигурация наиболее удаленного от карбонильной группы («концевого») асимметрического атома углерода, имеющего наибольший номер.
При совпадении конфигурации этого атома углерода с конфигурацией D-глицеринового альдегида моносахарид в целом относят к D-ряду. И наоборот, при совпадении с конфигурацией L-глицеринового альдегида считают, что моносахарид принадлежит к L-ряду. Знак вращения плоскости поляризации света моносахаридам непосредственно нельзя связать с их принадлежностью к D- или L-рядам, он определяется экспериментально. Так, среди альдопентоз и альдогексоз п-стереохимического ряда имеются как лево-так и правовращающие соединения. Углеводы могут существоватьтак же в циклической форме. Таутомерия – явление перехода открытых форм в циклическую. В растворах установление равновесия между четырьмя цикли-
ческими таутомерами моносахаридов протекает через открытую форму — оксоформу. Взаимопревращение а- и Р-аномеров друг в друга через промежуточную оксоформу называется аномеризацией. Таким образом, в растворе D-глюкоза существует в виде пяти таутомеров: а- и p-аномеров пиранозных и фуранозных цикличе- ских форм и оксоформы.
Ф
 ормулы
Хеуорса.
Для циклических форм моносахаридов
приняты перспективные формулы Хеуорса,
в которых циклы изображаются в виде
плоских многоугольников, лежащих
перпендикулярно плоскости рисунка.
Атом кислорода располагается в пиранозном
цикле в дальнем правом углу, в фуранозном
— за плоскостью цикла. Символы атомов
углерода в циклах не пишутся.
ормулы
Хеуорса.
Для циклических форм моносахаридов
приняты перспективные формулы Хеуорса,
в которых циклы изображаются в виде
плоских многоугольников, лежащих
перпендикулярно плоскости рисунка.
Атом кислорода располагается в пиранозном
цикле в дальнем правом углу, в фуранозном
— за плоскостью цикла. Символы атомов
углерода в циклах не пишутся.
Олигосахариды. В зависимости от количества остатков моносахаридов, входящих в молекулы олигосахаридов, последние делят па дисахариды, трисахариды и т. д. Наибольший интерес из олигосахаридов представляет группа дисахаридов—соединений, которые широко распространены в природе; многие из них имеют огромное практическое значение.
Сахароза представляет собой дисахарид типа глккозндо-гликозмда, составленный из остатков a-D-глюкопиранозы и β -D-фруктофуранозы. При гидролизе сахароза распадается на два моносахарида—a-D-глюкозу и β -D-фруктозу. Сахароза — один из самых распространенных в природе и практически наиболее важных дисахаридов. Она больше известна под названием тростникового или свекловичного сахара. Сахароза содержится также в соке многих других растений, но в сравнительно небольшой концентрации, высоко в соке клена, березы и пальмы.
Мальтоза относится к дисахаридам типа гликозндо-глкжоз и составлена из двух остатков a-D-глюкопиранозы, соединенных гликозидпой связью в положениях 1,4 Связь такого типа называют а-1,4-гликозидной связью. Мальтозу получают при гидролитическом расщеплении крахмала с помощью специфического фермента, которым очень богаты прорастающие зерна ячменя. Так как проросшие, а затем высушенные и измельченные семена ячменя называют солодом, а английское название солода—malt, то образующийся из крахмала при участии фермента солода дисахарид и получил наименование мальтозы.
Л актоза
является β-D-галактопиранозидо-4-D-глюкопиранозой,
содержится в молоке.
актоза
является β-D-галактопиранозидо-4-D-глюкопиранозой,
содержится в молоке.
Полисахариды. К полисахаридам относятся вещества, построенные из большого числа остатков моносахаридов или их производпых. Если полисахарид содержит остатки моносахарида одного вида, его называют гомополисахаридом. В том случае, когда полисахарид составлен из моносахаридов двух
видов или более, регулярно или нерегулярно чередующихся в молекуле, его относят к гетерополнсахаридам. К числу наиболее важных природных гомополисахаридов принадлежат
крахмал, гликоген (животный крахмал), клетчатка, декстран и хитин. Первые четыре полисахарида при гидролизе дают только D-глюкозу, последний—высвобождает производное D-ппокозы — N-ацетил-глюкозамин. Кроме того, известно много других гомополисахаридов. Так, полисахарид инулин при гидролизе дает только фруктозу, маннан—маннозу, галактан—галактозу, арабан—арабинозу и т. п. Названия этим полисахаридам даны по конечным продуктам их расщепления. К числу важнейших природных гетерополисахаридов относят гиалуроновую и хоидроитинсерную кислоты, гепарин, капсулярные полисахариды бактерий, агарозу, порфираны и каррагинаны красных водорослей, альгиновые кислоты бурых водорослей, структурные полисахариды простейших.
Биологическое значение полисахаридов разнообразно. Многие из них (крахмал, гликоген, инулин и др.) являются в растительных и животных организмах запасными питательными веществами. Некоторые полисахариды (например, хопдроитинссрпая кислота, капсулярные полисахариды и клетчатка) несут исключительно опорные и защитные функции. Ряд полисахаридов (маннаны, галактаны и др.) используется и как строительный, и как питательный материал. Тиалуроновая кислота, составляющая межклеточное вещество тканей животных, наряду со структурной функцией регулирует распределение жизненно необходимых веществ в тканях. Гепарин предотвращает свертывание крови в организме человека и животных. Во многих случаях полисахариды дают очень прочные комплексы с белками, образуя гликопротенны, выполняющие
в организме ряд ответственных функций. Крахмал — один из наиболее распространенных запасных полисахаридов растений. Он интенсивно накапливается в результате фотосинтеза и откладывается в семенах, клубнях и других частях растений. Глюкоза в составе крахмала находится в виде a-D-глюкопиранозы. Природный крахмал состоит из двух различных фракций, отличающихся по своему строению и свойствам. Примерно 20% крахмала составляет амилоза. Остальное приходится на вторую фракцию, получившую название амилопектина. Амилоза – линейного строения, амилопектин – разветвленного. Целлюлоза (клетчатка) растений имеет линейное строение, основной структурый элемент - β - D-глюкопираноза.
Обмен. Полисахариды и олигосахариды распадаются на простые соединения посредством реакций двух типов: 1. Реакции гидролиза (катализируют ферменты гидролазы) 2. Реакции фосфоролиза (катализируют ферменты фосфорилазы). Гидролазы – экзоферменты, которые катализируют гидролиз полисахаридов вне клеток или вне организма. В природе найдено несколько амилаз: a-амилаза (расщепляет a-1→4 связи в середине молекулы п/сах, образуя декстрины), ß-амилаза (отщепляет дисахариды мальтозы), мальтаза (расщепляет мальтозу), Υ-амилаза (последовательно отщепляет остатки глюкозы от невосстанавливающего конца молекулы крахмала), амило-1,6-глюкозидаза (расщепляет места разветвлений п/сах).
Другой путь распада полисахаридов и олигосахаридов — фосфоролиз. Фосфорилазы переводят полисахариды (гликоген) из запасной формы в метаболически активную форму. При действии фосфорилаз происходит разрыв гликозидной связи и перенос остатков глюкозы на молекулу фосфорной кислоты. В присутствии фосфорилазы гликоген распадается с образованием фосфорного эфира глюкозы (глюкозо-1-фосфата) без предварительного расщепления на более крупные обломки молекулы полисахарида. При этом образуется активированная форма глюкозы, энергетически богатая связь. При действии фосфорилаз происходит разрыв гликозидной связи и перенос остатков глюкозы на молекулу фосфорной кислоты.
В общей форме эту реакцию можно представить в виде:
(C6H10O5)n + H3PO4 –> (C6H10O5)n–1 + Глюкозо-1-фосфат
О бразовавшийся
в результате фосфоролитического распада
гликогена глюкозо-1-фосфат превращается
в глюкозо-6-фосфат под действием
фосфоглюкомутазы.
бразовавшийся
в результате фосфоролитического распада
гликогена глюкозо-1-фосфат превращается
в глюкозо-6-фосфат под действием
фосфоглюкомутазы.
Гликолиз - это последовательность реакций, приводящих к превращению глюкозы в пировиноградную кислоту (ПВК, пируват) с одновременным образованием ATФ.
П
 ервая
стадия:
ервая
стадия:
В дальнейший обмен вступает только 3-ФГА. В это соединение превращается и фосфодиоксиацетон, поэтому дальнейшим превращениям подвергаются 2 молекулы 3-ФГА. Образованием 3-ФГА (глицеральдегид-3-фосфата) как бы завершается первая стадия гликолиза.
Вторая стадия – наиболее сложная и важная. Она включает окислительно-восстановительную реакцию, сопряженную с субстратным фосфорилированием, в процессе которого образуется АТФ. Далее 3-ФГА в присутствии глицеральдегидфосфатдегидрогеназы, кофермента НАД и неорганического фосфата окисляется с образованием 1,3-дифосфоглицериновой кислоты (1,3-диФГК) и восстановленной формы НАД (НАДН). Далее 1,3-дифосфоглицериновая кислота превращается в 3-фосфоглицериновую кислоту, при этом образуется АТФ (процесс субстратного фосфорилирования). 3-ФГК в результате внутримолекулярного перемещения фосфатной группы (фермент – фосфоглицеромутаза) изомеризуется в 2-ФГК, который при участии енолазы теряет воду и превращается в енольную форму фосфопироградной кислоты (ФПК) – высокоэнергетического соединения. Далее образуется ПВК (пировиноградная кислота). В этой реакции также осуществляется субстратное фосфорилирование и образуется АТФ.
АТФ синтезируется при окислении 3-ФГА до 3-ФГК, протекающем через образование 1,3-диФГК, и при превращении 2-ФГК в фосфоенолпируват. Поскольку 2 молекулы АТФ потребляются в ходе первой стадии гликолиза в реакциях фосфорилирования, то общий итог гликолиза заключается в синтезе двух молекул АТФ. Нужно учесть и образование 2 НАДН+Н+
Г люконеогенез
(синтез «нового» сахара) – синтез глюкозы
из неуглеводных предшественников:
молочная и пировиноградная кислоты,
большинство аминокислот (гликогенные),
глицерол, промежуточные продукты цикла
трикарбоновых кислот. Глюконеогенез у
позвоночных наиболее интенсивно
протекает в клетках печени и почек (в
корковом веществе). Потребление алкоголя
резко тормозит глюконеогенез в печени,
вследствие чего понижается содержание
глюкозы в крови, возникает гипогликемия,
которая неблагоприятно сказывается на
функции мозга. Синтез полисахаридов
(глюканов) осуществляется путем переноса
моносахаридных остатков на
невосстанавливающий конец растущей
цепи полисахарида (реакции
трансгликозилирования) от соответствующих
субстратов: фосфорных эфиров моносахаридов,
уридиндифосфатсахаров и олигосахаридов.
Реакции трансгликозилирования ускоряются
ферментами - гликозилтрансферазами.
Для осуществления реакций
трансгликозилирования необходима
«затравка», т. е. наличие в реакционной
среде небольшого количества молекул
полисахарида, которые как бы предопределяют
тип связи, возникающий в процессе
синтеза.
люконеогенез
(синтез «нового» сахара) – синтез глюкозы
из неуглеводных предшественников:
молочная и пировиноградная кислоты,
большинство аминокислот (гликогенные),
глицерол, промежуточные продукты цикла
трикарбоновых кислот. Глюконеогенез у
позвоночных наиболее интенсивно
протекает в клетках печени и почек (в
корковом веществе). Потребление алкоголя
резко тормозит глюконеогенез в печени,
вследствие чего понижается содержание
глюкозы в крови, возникает гипогликемия,
которая неблагоприятно сказывается на
функции мозга. Синтез полисахаридов
(глюканов) осуществляется путем переноса
моносахаридных остатков на
невосстанавливающий конец растущей
цепи полисахарида (реакции
трансгликозилирования) от соответствующих
субстратов: фосфорных эфиров моносахаридов,
уридиндифосфатсахаров и олигосахаридов.
Реакции трансгликозилирования ускоряются
ферментами - гликозилтрансферазами.
Для осуществления реакций
трансгликозилирования необходима
«затравка», т. е. наличие в реакционной
среде небольшого количества молекул
полисахарида, которые как бы предопределяют
тип связи, возникающий в процессе
синтеза.