

Основы альгологии и микологии Охапкин
.pdfРис. 43. Схема строения клетки цианопрокариот на примере нитчатой водоросли Lyngbya: цм – цитоплазматическая мембрана; L1 – L4 – слои клеточной стенки; ч – чехол; фч – фибриллы чехла; по – пора в клеточной стенке; ц – цитоплазма; р – рибосомы; гв – газовые везикулы; г – гликогеноподобный полисахарид; т – тилакоиды; ф – фикобилисомы; нп – нуклеоплазма с нитями ДНК; цг – цианофициновые гранулы; в – волютин; с – септа
Вцентроплазме в виде гранул, палочек, сети или нитей локализована ДНК. Совокупность этих элементов представляет собой эквивалент ядра эукариот. В отличие от эукариот ДНК цианопрокариот не ограничена ядерной оболочкой и не связана с гистонами в структуры, подобные хромосомам.
Впериферической части клеток – хроматоплазме – локализованы тилакоиды – плоские мешочки, которые в поперечном сечении имеют вид пары параллельных мембран, более или менее плотно прилегающих
81
друг к другу и соединенных на концах. Тилакоиды, расположенные на периферии клетки, как правило, ориентированы параллельно клеточной стенке, но могут располагаться и перпендикулярно к ней. Иногда тилакоиды рассеяны по всей клетке. С тилакоидами связаны фотосинтетические пигменты. Добавочные пигменты синезеленых водорослей (фикоцианин, аллофикоцианин и фикоэритрин) в форме особых гранул – фикобилисом – локализованы на поверхности тилакоидов. В цитоплазме, не занятой тилакоидами и нуклеоплазмой (центром клетки, содержащим ДНК), находятся запасные вещества: гликогеноподобный полисахарид, зерна волютина (полифосфатные гранулы), цианофициновые (структурированные) гранулы, липидные включения и некоторые другие. Цианофициновые гранулы – немного угловатые мелкие тельца, состоящие из полимеров аминокислот аргинина и аспарагина, считают образованиями, специфическими для синезеленых водорослей и являющимися местом запаса азота. Рибосомы находятся как в центральной, так и в периферической частях клетки. Главный продукт ассимиляции – гликогеноподобный полисахарид. Хроматоплазма очень вязкая и в отличие от цитоплазмы клеток эукариот не находится в движении.
Клеточные оболочки синезеленых водорослей состоят в основном из пектиновых веществ (полисахариды) и часто ослизняются. Характерной их чертой является присутствие особого гетерополимера – муреина (пептидогликана), образующего в оболочке клеток синезеленых водорослей особый пептидогликановый слой, что сближает Cyanophyta с грамотрицательными бактериями и принципиально отличает их от эукариотических организмов. Клеточная стенка цианопрокариот многослойная, слои имеют разную структуру и плотность. Особое значение для поддержания формы клетки и прочности ее оболочки имеет муреиновый слой. Последний слой оболочки имеет структуру плазмалеммы – мембраны, окружающей протопласт. В продольных стенках клеток синезеленых водорослей и в септах, разделяющих соседние клетки, у нитчатых (гормогониевых) синезеленых водорослей имеются поры, через которые соединяются их цитоплазматические мембраны и протопласты. Эти цитоплазматические тяжи называются плазмодесмами. Для многих синезеленых водорослей характерно выделение большого количества слизи (сложные полисахариды), которая, по-видимому, является не непосредственным продуктом ослизнения оболочки, а чаще выделяется через оболочку из содержимого клетки.
Отличительной особенностью синезеленых водорослей является отсутствие вакуолей, наполненных клеточным соком. Однако у некоторых
82

из них постоянно или на определенной стадии развития образуются так называемые газовые вакуоли, или псевдовакуоли (рис. 44).
Газовые вакуоли, рассеянные |
|
|
по всей клетке или располагаю- |
|
|
щиеся у поперечных клеточных |
|
|
перегородок, состоят из тесно |
|
|
упакованных, наподобие сот, |
|
|
полых, одетых белковой мем- |
|
|
браной пузырей, |
заполненных |
|
газом. Они уменьшают удель- |
|
|
ный вес организма, способствуя |
|
|
его поднятию в верхние слои |
|
|
воды. Чаще других газовые ва- |
|
|
куоли встечаются у планктон- |
|
|
ных организмов. Они могут |
|
|
встречаться в клетках либо на |
|
|
протяжении всей |
вегетативной |
|
фазы развития водоросли, либо |
|
|
только на определенных стадиях |
|
|
развития. |
|
|
У ряда нитчатых синезеле- |
|
|
ных водорослей все клетки в |
|
|
нитях одинаковы, и такие тал- |
|
|
ломы называются гомоцитными. |
Рис. 44. Газовые вакуоли (а) |
|
У других нити состоят из веге- |
||
тативных клеток, |
гетероцист и |
в клетках Anabaena |
|
||
акинет, отличающихся по размерам и форме от вегетативных клеток. Такие талломы называются гетероцитными. Гормогониевые цианопрокариоты объединяют виды с нитчатой организацией строения тела. Нити свободные, очень редко срастающиеся с боков. Клетки тесно соединены друг с другом посредством тяжей цитоплазмы – плазмодесм и образуют трихомы. Трихом – совокупность клеток в пределах нити. Трихомы могут быть многорядными и часто окружены слизистым цилиндром – влагалищем. Трихом вместе с одевающим его влагалищем называют нитью. Некоторые представители образуют гетероцисты (рис. 45). Гетероцисты имеют сильно утолщенные стенки и толстую обвертку, образующуюся кнаружи от клеточной оболочки, благодаря чему выдерживаются механические воздействия. Клеточное содержимое гетероцист гомогенное, не содержит гранулярных включений (кроме рибо-
83

сом), волютина и цианофициновых зерен. В гетероцистах обнаруживаются хлорофилл и каротиноиды, но почти нет фикоцианина, аллофикоцианина и фикоэритрина. Роль гетероцист окончательно не выяснена. Обнаружено, что гетероцисты принимают участие в процессе фиксации атмосферного азота. Иногда гетероцисты могут прорастать, давая начало новым трихомам.
Рис. 45. Схема строения гетероцисты: цм – цитоплазматическая мембрана; L1 – L4 – слои клеточной стенки; пл – пластинчатый слой стенки гетероцисты; гс – гомогенный слой стенки гетероцисты; фс – фибриллярный слой стенки гетероцисты; мп – микроплазмодесмы; п – пора гетероцисты; с – септа гетероцисты; пр – пробка, замыкающая канал поры гетероцисты; м – мембраны гетероцисты; нд – рассеянные нити ДНК; р – рибосомы; вк – вегетативная клетка; гц – гетероциста
При ложном ветвлении во время разделения нитей трихомы чаще всего разрываются возле гетероцист, как бы ограничивая рост трихомов. Благодаря этому их раньше называли пограничными клетками. Нити с базальными и терминальными гетероцистами посредством их прикрепляются к субстрату. У некоторых видов акинеты располагаются рядом с
84

гетероцистой по одну или обе ее стороны. Подвижные жгутиковые стадии в жизненном цикле синезеленых водорослей отсутствуют.
Синезеленые водоросли размножаются главным образом вегетативно, реже бесполым путем, половое воспроизведение неизвестно. В основе размножения лежит деление клеток, которое у одноклеточных представителей является единственным способом их воспроизведения. Если клеточные деления быстро следуют друг за другом и получающиеся клетки не дорастают до размеров материнской, то образуются наноциты. Кроме того, размножение у нитчатых форм может происходить посредством гормогониев – отдельных участков трихомов, состоящих из молодых неспециализированных клеток и прорастающих в новые особи (рис. 46).
Рис. 46. Образование гормогониев у Lyngbya (слева) и акинет (а) у Anabaena (справа)
Споры (акинеты) одноклеточные, образуются путем дифференциации вегетативных клеток. Обычно они крупнее вегетативных клеток, по форме более или менее схожи с ними или отличаются от них. Акинеты покрыты плотными толстыми оболочками, состоящими из нескольких слоев. Благодаря толстым оболочкам и физиологическим изменениям в протопласте (накопление запасных веществ, увеличение количества цианофициновых зерен и ДНК, утрата газовых вакуолей), акинеты могут длительное время сохранять жизнеспособность в неблагоприятных условиях и при разнообразных сильных воздействиях (при низких и высоких температурах, при высыхании и сильном облучении). У некоторых цианопрокариот размножение происходит посредством мелких клеток, образующихся или внутри материнской клетки (эндоспоры –
85

беоциты) или отшнуровывающихся от верхушки материнской клетки (экзоспоры) (рис. 38). Типичного полового процесса у синезеленых нет, но у них обнаружены явления, при которых происходит частичное объединение геномов разных клеток. Отдел Cyanoprokaryota в современных системах содержит один класс Cyanophyceae и в зависимости от степени морфологической дифференциации делится по крайней мере на пять порядков, из которых ниже приводятся основные.
Порядок Chroococcales – Хроококковые – включает широко рас-
пространенные одноклеточные и колониальные свободноживущие или сидящие на субстрате водоросли. Колониальная слизь незаметная, чаще однородная. Клетки шаровидные, эллипсоидные, цилиндрические, реже другой формы. Размножение – делением клеток в одной или двух-трех плоскостях, включая образование наноцитов и почкование.
Рис. 47. Хроококковые цианопрокариоты: 1 – Synechocystis; 2 – Microcystis; 3 – Gloeocapsa
Род синехоцистис (Synechocystis) (рис. 47: 1) отличается шаровидными клетками, одиночными или после деления соединенными попарно, с тонкой оболочкой, не выделяющей видимой слизи.
Род микроцистис (Microcystis) (рис. 47: 2) имеет шаровидные клетки, у многих видов с газовыми вакуолями, беспорядочно расположенные в шаровидных, эллипсоидных или продырявленных колониях. Часто вызывает «цветение» воды, метаболиты некоторых видов ядовиты.
Род глеокапса (Gloeocapsa) (рис. 47: 3) характеризуется обычно шаровидными клетками. Клетки объединены в колонии, реже одиночные.
86
Колонии слизистые, состоящие из последовательно включенных друг в друга слизистых пузырей, из которых самые внутренние содержат клетки. Виды с обычно бесцветной слизью распространены в воде, другие преимущественно с окрашенной слизью (ярко-красная, желтая, фиолетовая и др.) – на суше: на влажных стенах, скалах, образуя на них налеты и корочки.
Род эндонема (Endonema) (рис. 38: 1) характеризуется короткими нитями, прикрепленными к субстрату базальной клеткой. Клетки нитей шаровидные или обратнояйцевидные. Конечная (базальная) клетка, как правило, цилиндрическая, по размерам больше, чем другие. Размножение преимущественно посредством эндоспор, образующихся в конечных клетках в числе 2–16. Эндоспоры большей частью выходят наружу, прикрепляются к субстрату и прорастают в новый таллом.
Род хамесифон (Chamaesiphon) (рис. 38: 2) имеет эллипсоидные, грушевидные или цилиндрические клетки, вокруг нижней части которых имеется чашевидная обертка – производное оболочки. Размножается экзоспорами, образующимися у части видов цепочками или понемногу. При разрастании образуются дисковидные скопления.
Порядок Oscillatoriales – Осциллаториевые. Трихомы всегда без гетероцист и в большинстве случаев без акинет, однорядные, с влагалищами или без них, неветвящиеся или образующие ложные, реже настоящие ветвления, по одному или помногу в каждом влагалище, одинаковой ширины на всем протяжении. Трихомы состоят из относительно одинаковых, не дифференцированных по форме и функциям клеток. Лишь конечные клетки могут иногда отличаться по форме и иметь утолщенные оболочки.
Виды рода осциллатория (Oscillatoria) (рис. 48: 1) часто образуют сине-зеленые пленки, покрывающие влажную землю, различные предметы, погруженные в воду, или плавают в виде толстых слизистых лепешек на поверхности стоячих водоемов. Нити осциллатории сложены из совершенно одинаковых цилиндрических клеток, за исключением верхушечных, которые по форме могут несколько отличаться от остальных. Нити обнаруживают своеобразное колебательное (осциллаторное) движение, сопровождающееся вращением нити вокруг собственной оси и ее поступательным движением. Размножается подвижными гормогониями, прорастающими в новые нити.
87
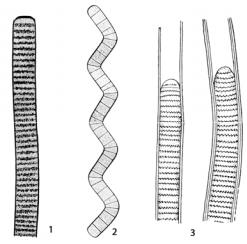
Рис. 48. Синезеленые водоросли порядка
Oscillatoriales:
1 – Oscillatoria;
2 – Spirulina;
3 – Lyngbya
Близок к осциллятории род спирулина (Spirulina) (рис. 48: 2), нити которой скручены в правильную спираль. Некоторые виды этого рода содержат много белка и с давних пор используются в пищу населением некоторых районов Африки. Спирулину культивируют с целью получения пищевых добавок, богатых протеинами, микроэлементами и другими биологически активными компонентами.
У представителей рода лингбия (Lyngbya) (рис. 48: 3) трихомы прямые, изогнутые, редко слабо спиралевидно или крючкообразно согнутые к концам, имеют одинаковую ширину на всем протяжении или суженные к обоим концам, у поперечных перегородок нередко с грануляциями, перешнурованные или не перешнурованные, обычно синезеленые или желто-зеленые, реже других оттенков. Нити одиночные или образующие дерновинки различной формы. Влагалища плотные, не склеивающиеся, бесцветные или желто-коричневые, тонкие или с возрастом толстые, иногда слоистые. Виды рода встречаются в стоячих и текучих водах, эпифитно на водных растениях, водорослях, раковинах моллюсков, эндофитно в слизи зеленых и синезеленых водорослей, а также в горячих источниках, в соленых и минеральных водах, почвах.
Порядок Nostocales – Ностоковые – объединяет водоросли с однорядными трихомами, постоянно с гетероцистами и нередко с акинетами, не ветвящиеся или ветвящиеся ложно, с хорошо оформленными или расплывающимися влагалищами.
88

Рис. 49. Цианопрокариоты порядка Nostocales:
Nostoc: 1 – внешний вид колоний; 2 – колония при большом увеличении микроскопа; 3 – Anabaena; 4 – Tolypothrix; Rivularia: 5 – колония при малом увеличении; 6 – часть колонии при большом увеличении
Род носток (Nostoc) (рис. 49: 1, 2) образует макроскопические, очень редко микроскопические колонии, постоянно более или менее шаровидные или реже эллипсоидные от 1 мм до 1 см и более (до величины куриного яйца) в поперечнике. Внутри колонии плотные или полые, мягкие или твердые, студенистые или иногда хрящеватые, почти всегда окружены плотным перидермом. Трихомы рыхло или тесно лежащие, большей частью радиально расположенные. Вегетативные клетки бочонкообразные, реже шаровидные или эллипсоидные. Гетероцисты шаровидные одиночные или иногда по несколько рядом, обычно крупнее вегетативных клеток. Акинеты образуются редко и для ряда видов совершенно неизвестны. Размножение посредством гормогониев. Большинство представителей обитает в воде. Самый крупный Nostoc pruniforme с шаровидными колониями и носток с колониями в виде слизи-
89
стых тяжей (N. flagelliforme), в которых нити расположены более или менее параллельными рядами, съедобны.
Представители рода анабена (Anabaena) (рис. 44; 46; 49: 3) имеют трихомы прямые или разнообразно (нередко спирально) изогнутые, одиночные или соединенные в мягкие слизистые, клочковидные или пленчатые дерновинки. Влагалища мягкие, расплывающиеся, очень редко ясно оформленные и заметные, обычно же вовсе не различимые в воде. Гетероцисты интеркалярные. Акинеты одиночные или располагаются рядами (цепочками), удалены от гетероцист или непосредственно примыкают к ним, располагаясь с одной или по обе стороны от них. Гетероцисты шаровидные, обычно с сильно отстающей наружной оболочкой. Акинеты шаровидные или эллипсоидные, одиночные или по две рядом, с крупнозернистым содержимым и утолщенной оболочкой. Род содержит около 100 видов, которые встречаются в воде и в наземных местообитаниях, среди них много возбудителей «цветения» воды. Наличие газовых вакуолей у некоторых видов постоянное, но в конце вегетационного периода они могут и исчезать. Отдельные виды часты на почве и наземных субстратах, многие фиксируют атмосферный азот.
Увидов рода толипотрикс (Tolypothrix) (рис. 49: 4) нити различно изогнутые, обильно растут в виде кустиков. Ложные ветвления одиночные, возникают у гетероцист. Влагалища крепкие, гомогенные или параллельно слоистые, бесцветные или желто-коричневые. Вегетативные клетки почти квадратные или длина их меньше или иногда больше ширины. Гетероцисты интеркалярные или чаще базальные (у основания нитей), нередко по несколько рядом. Гормогонии конечные. Акинеты наблюдаются очень редко, одиночные или располагаются цепочками. Ветви идут в том же направлении, что и основная нить, но обычно под некоторым углом к ней. Виды рода широко распространены в стоячих и текучих водах, на подводных растениях, створках раковин, среди других водорослей, многие сначала прикрепленные, затем свободно плавают, некоторые встречаются в почвах, на влажных скалах и камнях, на сырой земле, на стволах деревьев и на отмершей древесине.
Увидов рода ривулярия (Rivularia) (рис. 49: 5, 6) нити соединены в студенистые колонии более или менее правильно шаровидной или полушаровидной формы, прикрепленные или, реже, свободно плавающие. Внутри колонии нити располагаются радиально. Некоторые виды сильно инкрустированы известью. Нити и трихомы с базальными, реже также интеркалярными гетероцистами, к вершине утончаются, обычно переходят в волосок, неветвящиеся или ложно ветвящиеся, влагалища
90