

Основы альгологии и микологии Охапкин
.pdfУбольшинства водорослей пиреноиды погружены внутрь хлоропласта – их называют погруженными, у некоторых пиреноиды полупогруженные – расположены под оболочкой хлоропласта, немного выступая наружу. У некоторых видов эвгленовых, динофитовых, криптофитовых, золотистых, желтозеленых, диатомовых и бурых пиреноиды лежат за пределами хлоропласта, но тесно с ним связаны. Функция пиреноида заключается в концентрации рибулезобисфосфаткарбоксилазы с целью распределения ее между дочерними клетками и транспорта в места активного функционирования в хлоропласте. Наличие, размер пиреноида
иего обкладки во многом зависят от интенсивности освещения, температуры и других абиотических факторов. Обычно в хлоропласте находится один пиреноид, у некоторых водорослей до нескольких десятков. Чаще пиреноиды бывают округлой формы, иногда палочковидной или неправильной.
Уподвижных водорослей имеется фоторецепторная органелла – стигма или глазок (рис. 14). Функционально стигма связана с хлоропластом и жгутиковым аппаратом, поскольку она регулирует направление движения клеток путем улавливания световых импульсов, их трансформации и передачи жгутиковому аппарату. В световым микроскопе стигма представляет собой скопление кирпично-красного, оранжевого пигмента – астаксантина, растворенного в масле. Форма стигмы – сферическая, эллипсоидная, палочковидная, линзовидная или трапециевидная. У зеленых и криптофитовых стигма является частью хлоропласта и располагается на его периферии под оболочкой в передней части клетки. Она состоит из большого числа глобул, расположенных в 1–9 рядов.
Упредставителей золотистых, бурых и некоторых желтозеленых водорослей стигма также является частью хлоропласта. Она взаимно сориентирована со жгутиковым аппаратом. У этих водорослей при основании короткого жгутика формируется особое вздутие, так называемое парафлагеллярное или парабазальное тело, выполняющее рецепторную функцию. Оно располагается точно против вогнутой стигмы и отделяется от нее клеточным покровом и оболочкой хлоропласта. Стигма этого типа состоит из одного ряда пигментных глобул.
Уэвгленовых водорослей стигма не является частью хлоропласта, расположена за его пределами, но в непосредственной близости от жгутика. Жгутик несет утолщение – фоторецептор. Стигма состоит из нескольких десятков разных по размеру глобул, которые расположены рядами и обособлены друг от друга мембраной.
41

Рис. 14. Строение стигм различных водорослей: 1 – Chlamydomonas;
2 – Dinobryon; 3 – Euglena; 4 – Glenodinium; 5 – Nematodinium; а – хлоропласт;
б– стигма; в – пластинчатое тело; г – линзовидное тело;
д– ретиноид; е – пигментные тела; ж – парабазальное тело
42
Самые сложно устроенные стигмы наблюдаются у представителей динофитовых водорослей. У многих стигма расположена вне хлоропласта у основания жгутиков. Помимо двух рядов пигментных глобул, окруженных двухмембранной оболочкой, она включает своеобразное пластинчатое тело, которое связано с эндоплазматической сетью. У других видов стигма может достигать значительных размеров (20 мкм в длину и 15 мкм в ширину). Кроме пигментных глобул она включает линзовидное тело и ретиноид. Линзовидное тело фокусирует свет, ретиноид находится в его основании, ниже расположены пигментные глобулы. Благодаря такой организации стигмы свет попадает на глобулы концентированным пучком в преобразованном виде.
У некоторых жгутиковых (например, у некоторых видов рода Chlamydomonas, Dunaliella), а также подвижных стадий развития прикрепленных или пассивно плавающих водорослей стигмы нет и механизм фоторецепции неясен. У других водорослей, неподвижных в вегетативном состоянии, есть стигмы. В этом случае они рассматриваются как рудиментарные органеллы, утратившие свою функцию.
Во всех отделах эукариотических водорослей есть подвижные представители или стадии развития, например, зооспоры и гаметы. Все они обладают двигательным аппаратом. Движение осуществляется особыми структурами, которые могут быть временными и постоянными. К временным относятся ризоподии, или псевдоподии, к постоянным – аксоподии и жгутики, или ундулиподии. Ризоподиями называются длинные (до 140 мкм), тонкие, часто многочисленные выросты клетки, отграниченные от внешней среды плазмалеммой. Короткие, широко округлые временные выросты называются псевдоподиями. Аксоподии – это выросты клетки, внешне напоминающие щупальца и имеющие уплотненную осевую нить (рис. 15).
Жгутики (ундулиподии) – это длинные (до нескольких десятков мкм) или короткие выросты, характеризующиеся упорядоченной тонкой структурой, однотипной у всех водорослей. Жгутики, утратившие подвижность, называются псевдоцилиями (например, у тетраспоровых). У гаптофитовых водорослей (Haptophyta) кроме жгутиков есть еще короткий и более плотный и толстый направленный вперед вырост – гаптонема. С ее помощью клетки могут прикрепляться к субстрату, кроме того, она участвует в регуляции движения и процессе захвата клеткой пищевых частиц.
43
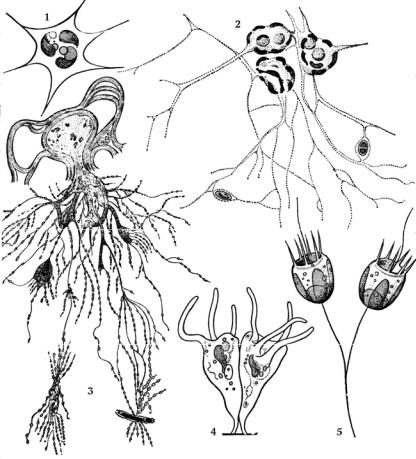
Рис. 15. Ризоподии, псевдоподии и аксоподии водорослей:
1–3 ризоподии: 1 – у Rhizochloris; 2 – у Chlorarachnion; 3 – у Chlamydomyxa; 4 – псевдоподии у Brehmiella; 5 – аксоподии и жгутики у Pedinella
Жгутики не обнаружены у цианопрокарит и прохлорофитовых, из эукариотических водорослей – у красных. Чаще количество жгутиков равно одному – двум, реже трем, четырем или восьми (рис. 16).
44

Г
Рис. 16. Основные типы строения жгутикового аппарата у монадных форм и стадий водорослей: 1 – Chlamydomonas; 2 – Carteria; 3 – Haematococcus; 4 – Tetraselmis; 5 – Pedinomonas; 6 – Micromonas; 7 – Oedogonium (зооспоры,
андроспоры, антерозоиды); 8 – Chara (антерозоиды); 9 – Euglena; 10 – Dinophyta; 11 – Cryptomonas; 12 – Chilomonas; 13 – Raphidophyta; 14 – Ochromonas;
15 – Chrysochromulina; 16 – Chromulina; 17 – Pavlova; 18 – Phaeophyta (зооспоры, гаметы); 19 – Bacillariophyta (антерозоиды); 20 – Eustigmatophyceae (зооспоры); 21 – Heterochloris; 22 – Vaucheria (антерозоид); 23 – Vaucheria (фрагмент зооспоры); 24 – Craspedomonadaceae (Monosiga); 1–4 – апикальное прик-
репление жгутиков; 8 – субапикальное; 18, 22 – латеральное; 10 – на брюшной стороне (вентральное); 4 – на дне «жгутиковой ямки»; 9 – на дне глотки; 11, 12 – в верхней части глотки; 1–4 – изоконтные, изоморфные, изодинамические жгутики; 13, 14, 18, 21, 22 – гетероконтные, гетероморфные, гетеродинамические; а
– акронематический жгутик (гладкий, с утонченной верхушкой); п – пантонематический (перистый, с двумя рядами мастигонем); с – стихонематический (гребенчатый, с одним рядом мастигонем); г – гаптонема
45

У зооспор вошерии (Vaucheria), дербезии (Derbesia) и эдогониума (Oedogonium) число жгутиков достигает нескольких десятков. По соотношению длины жгутиков в пределах одной клетки выделяют две группы водорослей – равножгутиковые (изоконтные) и разножгутиковые (гетероконтные). У последних жгутики занимают неодинаковое положение по отношению к клетке, имеют различия в частоте и интенсивности биений (гетеродинамичные). Длинный жгутик направлен вперед и работает энергичней болеекороткогозаднего, выполняющегорольруля.
Жгутики бывают гладкими или покрыты волосками – матигонемами
(рис. 17).
Рис. 17. Поверхностные структуры жгутиков:
1 – мастигонемы на жгутике сперматозоидов бурой водоросли Himanthalia;
2 – чешуйки на жгутиках сперматозоидов Chara
Мастигонемы расположены продольными параллельными рядами, одиночными или сдвоенными. Кроме матигонем, жгутики некоторых водорослей могут быть покрыты субмикроскопическими неминерализо-
46

ванными чешуйками, напоминающими чешуйки, расположенные на поверхности клеток. Тонкая структура жгутика у водорослей однотипна со строением жгутиков высших растений и животных. Снаружи жгутик покрыт мембраной, являющейся продолжением плазмалеммы клетки. Внутри жгутика находится матрикс, аналогичный матриксу клетки – гиалоплазма и система ориентированных микротрубочек, расположенных по схеме 9-9+2, которая называется аксонемой (рис. 18).
Рис. 18. Схема поперечного разреза жгутика эукариот (внизу), сложного жгутика динофитовых (1) и гаптонемы (2) гаптофитовых водорослей: а – жгутик в продольном сечении; б – жгутик на поперечном срезе; в – тело жгутика; г – стержнеподобная структура с поперечной исчерченностью; д – мастигонемы; е – микрофибриллы; ОЖ – оболочка жгутика; Д – наружные дублеты, образованные А- и В-микротрубоч- ками; Р – боковые ручки из динеина; ЦМ – центральные микротрубочки; ЦФ – центральный футляр; ДС – интердублетные соединения из нексина; РС – радиальныеспицы
Аксонема – это сложный комплекс микротрубочек, состоящих из белка тубулина, связанного с некоторыми другими белками. Изгибанием аксонемы обусловлено биение жгута. Аксонема состоит из девяти пар периферических микротрубочек и двух центральных. Центральная пара микротрубочек заключена в центральную капсулу, образованную
47

тонкими загнутыми белковыми отростками, прикрепленными к микротрубочкам. Периферические микротрубочки составляют пары, которые называются дублетами. Каждый дублет образован микротрубочкой А и микротрубочкой В, первая состоит из тринадцати субъединиц тубулина, вторая – из десяти. Микротрубочка А несет короткие динеиновые ручки
– выросты из белка динеина, направленные к соседнему дублету. Динеиновые ручки необходимы для изгибания жгутиков. Соседние дублеты микротрубочек связаны друг с другом эластическими связями, образованными белком нексином. Этими связями аксонема жгутика оказывается скрепленной как бы обручем. От каждого дублета внутрь аксонемы отходит радиальная спица, заканчивающаяся глобулярным образованием вблизи центральной капсулы.
В основании каждого жгутика, вблизи поверхности клетки находится базальное тело жгутика, или кинетосома, представляющая собой короткий (до 2 мкм длиной) цилиндр. По структуре кинетосомы идентичны центриолям и во многих случаях могут превращаться друг в друга. Центриоли обычно представлены парами, члены которых расположены под прямым углом друг к другу. Каждая центриоль состоит из девяти триплетов (состоящих из трех) микротрубочек (рис. 19).
Новая пара центриолей образуется по соседству с уже существующей. Этот процесс происходит сходно у всех эукариот. Функции центриолей – образование жгутиков и веретена деления, формирующегося при митозе и мейозе.
Стенки базального тела, периферическая его часть образована девятью триплетами микротрубочек, внешний диаметр базального тела равен диаметру аксонемы. Система микротрубочек базального тела составляет единое продолжение системы микротрубочек аксонемы, но в базальном теле к продолжению пары микротрубочек аксонемы при-
48

соединяется третья неполная микротрубочка. Центральная пара микротрубочек аксонемы оканчивается выше базального тела, у основания жгутика, и поэтому центр базального тела лишенмикротрубочек.
От базального тела отходят микротрубочковые корни жгутиков, преимущественно состоящие из 2–4 микротрубочек, которые у большинства водорослейрасположенысимметрично и образуют характерные крестообразныефигуры(рис. 20).
Базальные тела жгутиков связаны друг с другом особыми фибриллярными структурами – связывающими тяжами. Кроме микротрубочковых корней от базальных тел могут отходить фибриллярные корни или один крупный поперечно исчерченный фибриллярный тяж – ризопласт. Число, расположение и структура связывающих тяжей и фибриллярных корней сильно варьируют у разных водорослей. У части зеленых и водорослей стреп-
тофитов корневая система имеет асимметричное строение, связанное с наличием так называемой многослойной структуры. Она имеет очень сложное строение и сходство в организации с аналогичной структурой в сперматозоидах высших растений.
Корни жгутиков направлены на периферию клетки, выполняют роль цитоскелетных образований. Фибриллярный тяж подходит к ядру, в зону расположения центриолей, осуществляя связь жгутикового аппарата с ядром.
49

|
У гаптофитовых водорослей помимо |
|
жгутиков есть особое образование – гап- |
|
тонема, имеющая вид направленного впе- |
|
ред жгутикоподобного выроста (рис. 21). |
|
Гаптонема состоит из микротрубочек, |
|
расположенных по кругу и окруженных |
|
частично или полностью эндоплазматиче- |
|
ским ретикулумом (рис. 18: 2). Это обра- |
|
зование принимает участие в движении |
|
клеток, а также в процессе улавливания и |
|
заглатывания клеткой твердых органиче- |
|
скихчастиц. |
|
Митохондрии у водорослей – крупные, |
|
овальные тельца, окруженные оболочкой |
|
из двух мембран. Их число, размеры и |
Рис. 21. Плавающие клетки |
форма варьируют в широких пределах в |
зависимости от возраста клетки и ее |
|
гаптофитовой водоросли |
функционального состояния. |
Chrysochromulina: |
Клетки большинства водорослей со- |
г – гаптонема; жг – жгутик |
держат одно ядро, но есть виды, содер- |
|
жащие в клетках 2–3 и более ядер, до |
нескольких десятков и сотен, иногда до нескольких тысяч (например, в старых клетках красной водоросли гриффитзии). Размеры ядра колеблются от 1 до 45 мкм, составляя в среднем около 4 мкм. Ядра окружены двухслойной оболочкой, пронизанной порами, внутри которой находится нуклеоплазма, одно или несколько ядрышек и хроматин. Хроматин в виде скрученных глыбок и нитей обычно располагается вдоль ядерной оболочки. Хроматин – это форма существования хромосом в интерфазном ядре. Хромосомы обычно имеют центромеры и перетяжки и претерпевают циклические преобразования, находясь то в спирализованном состоянии, то деспирализуются.
Степень спирализации хромосом, соотношение объема нуклеоплазмы и ядрышек в интерфазном ядре у разных водорослей сильно варьирует. Наряду с типичными картинами митоза у водорослей выявлены многообразные его разновидности, связанные с различной выраженностью веретена деления, наличием или отсутствием центриолей, поведением ядерной оболочки и ядрышек. Выделяют закрытый митоз, когда ядерная оболочка в период митоза остается непрерывной (у некоторых представителей зеленых, эвгленовых, бурых, желтозеленых и красных) (рис. 22).
50