

Основы альгологии и микологии Охапкин
.pdfды легко вступают в симбиоз, например Tetraselmis с морским плоским червем Convoluta.
Виды рода пирамимонас (Pyramimonas) (рис. 112) встречаются как в морях, так и в пресных водах.
Рис. 112. Внешний вид (слева) и детали строения клетки (справа) Pyramimonas: А – схема строения клетки; Б – строение чешуек: 1–3 – чешуйки тела
(1 – внутренняя, 2 – средняя, 3 – наружная); 4, 5 – чешуйки жгутика, внутренняя
(4) и наружная (5); нчт – наружная чешуйка тела; вчт – внутренняя чешуйка тела; счт – средняя чешуйка тела; ж – жгутик; вчж – внутренняя чешуйка жгутика; нчж – наружная чешуйка жгутика; бт – базальные тела; жк – жгутиковый корень; рч – резервуар чешуек; д – диктиосома; я – ядро; х – хлоропласт; п – пиреноид; с – стигма; м – митохондрия; т – трихоциста
Клетки имеют обратнопирамидальную, обратногрушевидную или овальную форму; в передней части в поперечном сечении клетки квадратные. Кнаружи от плазматической мембраны клетки покрыты 2–5 слоями чешуек с очень характерной для каждого вида структурой. Четыре жгутика выходят из ямки, расположенной на переднем конце клет-
211
ки, и покрыты двумя слоями чешуек. Помимо чешуек, жгутики несут волосовидные отростки. Чешуйки всех типов формируются в цистернах аппарата Гольджи, затем они транспортируются к специальному резервуару чешуек, который специальным протоком открывается на поверхность клетки. Чешуйки имеют органическую природу, фибриллярную структуру, но их химический состав изучен недостаточно. Основания четырех жгутиков связаны поперечно-исчерченной структурой и соединяются с пиреноидом посредством ризопласта.
Ядро одно, занимает в клетке боковое положение. Хлоропласт разделен продольно на 4, 8 или 12 лопастей, которые простираются почти до его основания. Пиреноид один в задней части хлоропласта, окружен крахмальными зернами. Ламеллы хлоропласта сформированы из 2–5 тилакоидов; стигма является частью хлоропласта и состоит из 2–4 рядов липидных глобул. В клетке имеются трихоцисты, выбрасывающиеся наружу в виде нитей. Размножение осуществляется путем продольного деления клетки. Известен половой процесс по типу хологамии. Зигота прорастает четырьмя зооспорами, сходными с взрослыми клетками пирамимонаса, но меньших размеров.
Среди коккоидных представителей празиновых широко распространена в планктоне морей водоросль галосфера (Halosphaera). Неподвижная стадия этой водоросли представлена шаровидными вегетативными клетками с оболочкой, которые часто рассматриваются как цисты. Подвижная фаза – это четырехжгутиковые монадные клетки, сходные с пирамимонасом и способные размножаться путем продольного деления. Вегетативные клетки (цисты) могут заметно увеличиваться в размерах и в них образуются подвижные монадные клетки. После их высвобождения из цист эти клетки двигаются, во время периода движения размножаются путем деления, затем округляются, теряют жгутики и превращаются в неподвижные одетые оболочкой клетки (цисты).
Положение празинофитовых водорослей в системе зеленых водорослей спорно, ранее их включали в состав порядка Volvocales и только в последние десятилетия, после исследований их тонкой ультраструктуры, предложено рассматривать эти водоросли в качестве самостоятельного класса. Все больше сторонников находит точка зрения, что представители этой группы являются прообразом предков или близких к ним форм, давших начало различным классам зеленых водорослей.
У последующих классов зеленых водорослей жгутики, прикрепленные крестообразно, базальные тела расположены по типу 11–5, у представителей класса Chlorophyceae – по типу 1–7 или 12–6.
212
Класс Ulvophyceae – Ульвофициевые водоросли – включает водо-
росли коккоидной, нитчатой, паренхиматозной и сифонокладальной структуры. Жгутиковый аппарат монадных стадий организован по типу 11–5 с четким сдвигом базальных тел относительно друг друга против часовой стрелки. Образование клеточной стенки при цитокинезе происходит по типу борозды без участия фикопласта. Бесполое размножение осуществляется зооспорами или апланоспорами, половое – изогамия или гетерогамия.
Представители порядка Ulotrichales (Codiolales) – Улотриксо-
вые (Кодиоловые) – могут быть одноклеточными или имеют талломы в виде неразветвленной или ветвящейся нити, сложенной из одноядерных клеток, реже таллом пластинчатый, но в онтогенезе развивающийся из нитчатого. Водоросли обычно микроскопические или различимые невооруженным глазом, нередко довольно крупные, достигающие в длину нескольких дециметров. Хлоропласт пластинчатый, пристенный. Бесполое размножение зооспорами или апланоспорами, половой процесс – изогамия и гетерогамия, но характерен гетероталлизм. В жизненном цикле имеется одноклеточная кодио- лум-стадия, при прорастании которой происходит мейоз с образованием гаплоидных зооспор.
Широко распространены в пресных и морских водах виды рода улотрикс (Ulothrix), чаще других встречается Ulothrix zonata (рис. 30; 113).
Таллом улотрикса имеет вид неветвящейся нити, состоящей из одного ряда клеток цилиндрической формы и одетых толстой оболочкой. Под оболочкой находится постенная цитоплазма, один пластинчатый хлоропласт, имеющий форму незамкнутого кольца и содержащий несколько пиреноидов. Клетки улотрикса одноядерные, центр клетки занят вакуолью клеточного сока. Нить прикрепляется к субстрату клиновидной клеткой (рис. 30), которая служит только для прикрепления и лишена хлоропластов и пигментов (бесцветна). Все остальные клетки несут функции вегетации и размножения. Нарастание таллома происходит за счет вегетативного клеточного деления любых клеток в пределах нити в течение вегетативной фазы жизненного цикла.
213
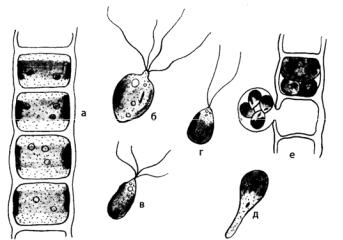
Рис. 113. Ulothrix zonata: а – внешний вид нити; б, в – зооспоры; г – гамета; д – прорастание зооспоры; е – образование зооспор
При бесполом размножении в клетке образуется от одной до шестнадцати, реже тридцати двух четырехжгутиковых зооспор. Процесс образования зооспор происходит в направлении от апикальной клетки нити к клеткам, расположенным в ее основании. Зрелые зооспоры, заключенные в слизистой обвертке, высвобождаются из материнской клетки через отверстие в клеточной стенке.
Различают микро- и макрозооспоры, отличающиеся размерами, положением глазка и временем плавания. После периода плавания зооспора останавливается, сбрасывает жгутики, прикрепляется боком к субстрату и прорастает в нить. При прорастании зооспоры происходит дифференциация ее протопласта на две части: нижнюю, лишенную хлоропласта, превращающуюся в базальную клетку – ризоид, и верхнюю, которая при дальнейших делениях дает вегетативные клетки нити. Иногда вместо зооспор образуются толстостенные апланоспоры. Перед окончанием вегетативного роста или при наступлении неблагоприятных условий растение переходит к половому размножению.
При половом размножении в клетках в числе 8 (4) – 32 (64) образуются двужгутиковые изогаметы. Гаметы выходят из клетки, заключенные в слизистый пузырь, в котором они двигаются, пока слизь не рас-
214

плывется в воде. У Ulothrix zonata наблюдается гетероталлизм, т.е. гаметы сливаются из разных нитей, различающихся по половому знаку («+» и «–»). После копуляции образуется четырехжгутиковая планозигота, которая после периода плавания прикрепляется к субстрату, округляется, втягивает жгутики и прорастает в одноклеточный диплоидный спорофит (Codiolum-стадия). Последний имеет обычно грушевидную или сферическую форму и плотную оболочку. После короткого или длительного периода покоя протопласт кодиолум (спорофита) начинает делиться с редукцией числа хромосом. В результате образуется 4–16 четырехжгутиковых зооспор. Зооспоры оседают на субстрат и прорастают в нить, так же, как и зооспоры, возникающие при бесполом размножении. Если условия для бесполого и полового размножения неблагоприятны, улотрикс может размножаться путем фрагментации нитей. При наступлении неблагоприятных условий улотрикс может переходить в пальмеллевидное состояние. При этом клетки округляются, их стенки ослизняются, нить распадается на отдельные фрагменты и клетки. В таком состоянии клетки могут делиться. При наступлении благоприятных условий каждая клетка такой пальмеллевидной стадии может превращаться в зооспору. Виды улотрикса являются одними из самых широко распространенных ничаток, развивающихся в бентосе стоячих и слабо проточных вод.
Цикл развития Ulothrix zonata
У видов рода монострома (Monostroma) гаметофит пластинчатый однослойный, размером от нескольких сантиметров до одного метра, прикрепляющийся к камням дисковидной подошвой (рис. 114).
215
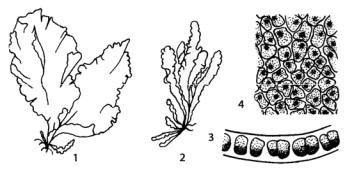
Рис. 114. Monostroma: 1, 2 – общий вид слоевища;
3 – поперечный срез слоевища; 4 – вид клеток с поверхности слоевища в средней части
Спорофит (стадия Codiolum) у моностромы одноклеточный. Например, у Monostroma grevillei макроскопическое слоевище гаметофита на ранних стадиях развития имеет мешковидную форму, которая по мере роста таллома преобразуется в однослойную пластинку. Гаметы, образующиеся на гаметофите, выходят в воду через отверстие во внешней оболочке клеток и сливаются с образованием зиготы. Зигота развивается в одноклеточный диплоидный спорофит. При этом она сильно разрастается и покрывается толстой оболочкой. Через некоторое время протопласт спорофита делится с образованием гаплоидных зооспор. Таким образом, в данном случае имеется гетероморфная смена форм развития. Встречается водоросль в литорали и сублиторали морей, может выдерживать опреснение.
Порядок Ulvales – Ульвовые – включает морские и пресноводные водоросли-макрофиты с талломами, состоящими из одного-двух слоев одноядерных клеток. Жизненный цикл гапло-диплофазный с изоморфной сменой форм развития. Половой процесс – изоили гетерогамия, гаметы двужгутиковые, зооспоры – четырехжгутиковые.
Это водоросли с пластинчатым паренхиматозным листовидным талломом без полости, или пузыревидные и трубчатые с однослойными стенками и полостью в середине таллома. Все эти типы талломов развиваются из прикрепленной к субстрату нити, построенной по типу улотрикса.
216
После ряда поперечных делений в такой однорядной нити клетки начинают делиться также и продольно, в результате чего образуются пластинчатые талломы.
У видов рода ульва (Ulva) (рис. 33: 2; 115), широко известных как зеленый морской салат, талломы имеют вид пластинок светлозеленого цвета с гофрированными краями, прикрепленных к субстрату суженным в короткий черенок основанием.
Это одна из массовых водорослей, обитающих в морях и солоноватых водах. Клетки одноядерные с постенным хлоропластом, имеющим вид не замкнутой в кольцо пластинки. У
ульвы наблюдается изоморфная смена форм развития: полового поколения – гаметофита и бесполого поколения – спорофита. Бесполые растения – спорофиты диплоидны. При образовании на них зооспор диплоидное ядро делится с мейозом, и гаплоидные зооспоры прорастают в гаплоидные растения – гаметофиты, на которых формируются гаметы. Половой процесс – изогамия или гетерогамия. У ульвы наблюдается гетероталлизм, копулируют гаметы с разными половыми знаками. Диплоидная зигота без периода покоя прорастает в диплоидное растение – спорофит. Оба поколения различаются только цитологически, и по органам размножения, которые на них развиваются, внешне они неотличимы. Такая смена форм развития получила название изоморфной. У некоторых видов отмечено партеногенетическое развитие гамет в новую особь.
Ульва обитает в литорали и сублиторали северных и южных морей на камнях, скалах, раковинах; может выносить значительное опреснение. Водоросль богата витаминами В и С и используется как приправа к различным блюдам.
217

Цикл развития Ulva lactuca
|
У энтероморфы (Enteromorpha) (рис. 116) |
|
трубчатые талломы с однослойной стенкой, |
|
крупные, разнообразной формы, часто вет- |
|
вящиеся. |
|
Русское название этой водоросли – ки- |
|
шечница – отражает сходство ее таллома с |
|
кишкой или трубкой. Растет на различных |
|
субстратах у уреза воды и глубже или сво- |
|
бодно плавает в соленых, солоноватых и |
|
пресных, часто загрязненных водах. Гамето- |
|
фит и спорофит внешне сходны и различа- |
|
ются только во время размножения. Зооспо- |
|
ры выходят в воду через округлое отверстие |
|
во внешней оболочке клеток спорофита и |
|
прорастают путем последовательных деле- |
Рис. 116. Внешний |
ний в однорядную нить. Затем из нити фор- |
вид и поперечный |
мируется двухслойная пластинка, слои кото- |
срез молодого |
рой впоследствии разъединяются, давая |
участка таллома |
трубчатое слоевище гаметофита, ничем |
Enteromorpha |
внешне не отличающееся от слоевища спо- |
|
рофита. Таким образом, у энтероморфы, как |
и ульвы, отмечается изоморфная смена форм развития со спорическим мейозом.
К классу Trebouxiophyceae – Требуксиофициевые – относятся преимущественно одноклеточные коккоидные, реже сарциноидные и разветвлено-нитчатые водоросли. Жгутиковый аппарат репродуктивных клеток имеет крестообразные микротрубочковые корешки и базальные
218
тела, сдвинутые против часовой стрелки по типу 11–5. Дочерние клетки возникают внутри материнской по типу схизогонии, в чем проявляется сходство с водорослями порядка Chlorococcales класса Chlorophyceae. В отличие от последних, у Trebouxiophyceae при митозе центриоли располагаются по бокам веретена (метацентрическое веретено). Размножение вегетативное, бесполое (зооспоры, автоспоры), половое.
Виды рода требуксия (Trebouxia) отличаются сферическими клетками с осевым звездчатым хлоропластом с пиреноидом. Бесполое размножение осуществляется голыми зооспорами. Является одним из распространенных фикобионтов в лишайниках, кроме того, встречается в свободном виде во вневодных местообитаниях, например, на коре деревьев.
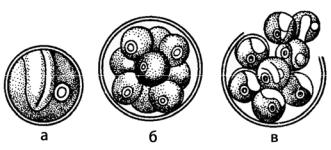
Среди требуксиофициевых водорослей, при бесполом размножении которых образуются автоспоры, наиболее известна хлорелла (Chlorella) (рис. 117).
Рис. 117. Chlorella: а – вегетативная клетка; б, в – образование и выход автоспор
Виды этого рода широко распространены в пресных и солоноватоводных водоемах, на сырой земле, коре деревьев и других влажных субстратах. Это мелкие шаровидные или эллипсоидные клетки, одетые гладкой слоистой оболочкой, содержат один обычно чашевидный хлоропласт с пиреноидом и одно ядро. Наружные слои оболочки клетки пропитаны спорополленином – продуктом полимеризации каротиноидов, придающим оболочке устойчивость к воздействию факторов внешней среды, в том числе и к химическим агентам (ферментам, кислотам, щелочам). При бесполом размножении протопласт клетки образует 4–8, реже больше автоспор, которые внутри материнской клетки одеваются собственными оболочками и принимают форму материнского организ-
219
ма. Автоспоры высвобождаются наружу после разрыва оболочки материнской клетки. Один из видов этого рода легко вступает в симбиотические отношения с простейшими (Paramecium bursaria). За счет фотосинтеза простейшее получает органическое вещества, а водоросли – продукты обмена хозяина. Многие виды относятся к аэрофитным, т.е. обитают в наземно-воздушной среде, а не в воде. Хлорелла является объектом культивирования, поскольку сухая биомасса водоросли содержит много белков, улеводов, витаминов, микроэлементов и других физиологически активных веществ. Ее культивируют в Японии, Канаде, США, Франции, Новой Зеландии, Австралии, Корее и других странах. Культивирование хлореллы в замкнутых экосистемах космических кораблей позволило подойти к проблеме снабжения космонавтов кислородом и компонентами пищевого рациона в период продолжительных полетов. Лабораторные культуры этой водоросли являются удобным объектом для выяснения физиологических механизмов фотосинтеза и дыхания, биологического саморегулирования, биосинтеза различных соединений, для решения фундаментальных проблем физиологии, биохимии, генетики и других наук.
Представители класса Cladophorophyceae – Кладофорофициевые
– имеют исключительно сифонокладальную структуру таллома, имеющего вид разветвленных кустиков или слабо ветвящихся нитей, образованных многоядерными клетками.
Порядок Cladophorales (Siphonocladales) – Кладофоровые (Сифо-
нокладовые) водоросли – это макроскопические, нитчатые, простые или разветвленные, прикрепленные к субстрату или свободно плавающие в толще воды талломы сифонокладальной структуры. Слоевище образовано многоядерными клетками или сегментами, образующимися при особом типе клеточного деления (сегрегативное клеточное деление), при котором процесс деления ядер не сопровождается образованием клеточных перегородок, а образование отдельных сегментов происходит независимо от кариокинеза (рис. 23: 3). Макроскопические талломы прикрепляются к субстрату неклеточными или сегментированными ризоидами. Хлоропластная система состоит из многочисленных дисковидных или многоугольных участков, соединенных тонкими тяжами, и в световом микроскопе хлоропласт имеет вид сетчато продырявленной пластинки. Оболочка сифонокладовых водорослей нередко пропитана солями кальция, магния, кремния, иногда наблюдаются вкрапления солей железа. В целом оболочка отличается большой прочностью, что определяет стабильную, весьма разнообразную форму слоевищ.
220