

Основы альгологии и микологии Охапкин
.pdfкороткий вторичный стебелек. Всего на талломе образуется до 40 пластин. Растет нереоцистис на глубине до 20 м, при этом верхняя часть таллома плавает на поверхности воды. Распространен в основном у Тихоокеанского побережья Северной Америки.
Алария (Alaria) (рис. 107: 4) имеет пластинку с продольным ребром, идущим от ствола до вершины. Края пластины ровные или с длинными боковыми лопастями. По бокам стволика имеются пластинки на тонких стебельках (спорофиллы), на которых развиваются одноклеточные спорангии. Спорофиллы скученные, длинноланцетовидной формы, до 25 см длиной и 1–3 см шириной. Слоевище достигает в длину 2–3 м и прикрепляется довольно слабо разветвленными ризоидами. Все виды рода распространены в нижней литорали и сублиторали морей Северного полушария и предпочитают места с постоянным движением воды. В России встречается по побережью Белого, Баренцевого и Карского морей.
Класс Cyclosporophyceae – Циклоспоровые включает водоросли с крупными, сложно дифференцированными в анатомическом и морфологическом отношениях талломами. У представителей класса гаметофит утратил способность к самостоятельному существованию и представляет собой слой, выстилающий особые углубления (концептакулы или скафидии), развивающиеся на спорофите. У всех циклоспоровых мейоз происходит при образовании гамет. Процесс оплодотворения и развитие зиготы происходит вне растения. Класс делится на три порядка, наиболее крупным из которых является порядок фукусовые – Fucales.
Порядок Fucales – Фукусовые. Растения в большинстве случаев многолетние. Слоевище кустистое, разнообразной величины и формы, большей частью сложного морфологического строения, рассеченное на основную цилиндрическую или пластинчато-лентовидную часть, короткий стволик и конусовидную или дисковидную подошву. Рост осуществляется посредством одной – немногих верхушечных клеток. Бесполого размножения нет, половой процесс – оогамия. Антеридии и оогонии развиваются в специальных обособленных участках, погруженных в слоевище – скафидиях. Скафидии разбросаны по всему слоевищу или сконцентрированы в определенных местах, в специальных образованиях
– рецептакулах. Оогонии с 1–8 яйцеклетками. Размеры талломов фукусовых меньше, чем у ламинариевых, но они могут достигать 10 м длины. Широко распространены в северных широтах, где образуют заросли в зоне прилива – отлива (литорали) и сублиторали.
201
Виды рода фукус (Fucus) (рис. 108) достигают 0,5–1,0 м в длину, реже более, и имеют талломы в виде сильно дихотомически разветвленных кустиков.
Нижняя часть плоского таллома переходит в цилиндрический черешок, прикрепленный к субстрату с помощью диска. У части видов по бокам от средней жилки расположены воздушные пузыри, образующиеся в результате разрыва тканей внутри слоевища и поддерживающие таллом в вертикальном положении в воде. На концах зрелого таллома, которые вздуваются, принимают более светлую желто-оранжевую окраску и превращаются в рецептакулы, формируются концептакулы или скафидии. Рецептакулы округлой или овальной формы, с тупыми верхушками. Скафидий – это углубление в талломе диплоидного растения, дно которого выстлано клетками, образующимися за счет деления особой клетки – проспоры, или инициальной клетки скафидия.
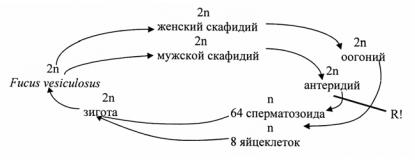
Клетки выстилающего слоя в верхней части зрелого концептакула развивают волоски, выступающие из его отверстия. Из клеток дна скафидия образуются разветвленные (в мужских скафидиях) или неразветвленные (в женских скафидиях) многоклеточные нити – парафизы. У фукусовых есть однодомные и двудомные виды. У однодомных оогонии и антеридии могут развиваться в одном скафидии. Антеридии образуются на концах разветвленных парафиз и при созревании дают 64 сперматозоида. Сперматозоиды гетероконтные и гетероморфные, в отличие от монадных клеток остальных бурых водорослей задний жгутик длиннее переднего. Оогонии прикреплены к стенке концептакула клеткой – ножкой.
Деление первичного ядра, как и в антеридиях, редукционное, в оогонии фукуса образуется 8 яйцеклеток. Стенка оогония трехслойная, при созревании наружный слой разрушается и зрелые яйцеклетки, окруженные средним и внутренним слоями оогония, выдвигаются наружу через отверстие скафидия. Затем разрушается средний слой и последним в воде – внутренний, после чего яйцеклетки выходят в воду, где и оплодотворяются сперматозоидами. У однодомных видов с обоеполыми скафидиями яйцеклетки оплодотворяются в самом концептакуле и уже оплодотворенные высвобождаются в воду. У Fucus serratus была установлена химическая структура вещества, хемотаксически привлекающего сперматозоиды к яйцеклетке. Это вещество оказалось низкомолекулярным углеводородом и было названо фукосерратеном (С8Н12).
202

Рис. 108. Строение таллома и размножение у Fucus: a – общий вид таллома; б – разрез женского скафидия; в – разрез мужского скафидия; г – оогоний;
д, е – выход яйцеклеток из оогония; ж – веточка
сантеридиями; з – выход сперматозоидов из антеридиев;
и– сперматозоид
203
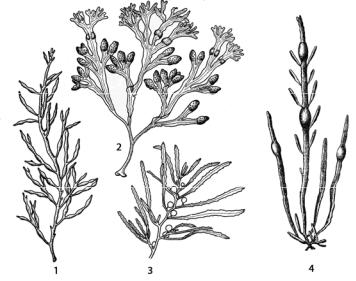
Рис. 109. Бурые водоросли порядка Fucales: 1 – Cystoseira barbata; 2 – Fucus vesiculosus; 3 – Sargassum; 4 – Ascophyllum nodosum
Цистозейра бородатая (Cystoseira barbata) (рис. 109: 1) растет на каменистых и каменисто-ракушечных грунтах в Черном и Азовском морях на глубине до двадцати метров. От прикрепительного диска, как правило, отходит один высокий «ствол» с поочередно или почти беспорядочно расположенными боковыми «ветвями». Вторичные веточки многократно ветвящиеся, цилиндрические, короче, чем основные, и часто собраны метелками. Воздушные пузыри одиночные или в цепочках, чаще располагаются на конечных веточках. Рецептакулы обоеполые, развиваются зимой и отчасти летом; в оогонии одна яйклетка. Светолюбива, богата альгиновой кислотой.
У фукуса пузырчатого (Fucus vesiculosus) (рис. 109: 2) талломы от 15 до 150 см высотой, оливково-бурые в верхней части и почти черные в основании. Края ветвей ровные. Водоросль живет в литоральной зоне и очень плотно прикрепляется к камням конической подошвой. Слоевища имеют округлые или овальные воздушные пузыри, попарно рас-
204
полагающиеся по бокам ребра. Пузыри не образуются на талломах, развивающихся в местах прибоя. Рецептакулы круглые или овальные, простые или вильчато разветвленные. Имеются мужские и женские экземпляры, отличающиеся размерами талломов. Мужские талломы нередко мельче женских, растущих рядом, с более узкими ветвями и более мелкими рецептакулами. Наиболее пышные заросли этот вид образует на литорали в полузащищенных местах. У берегов России встречается в Белом, Баренцевом, Карском и Балтийском морях.
Цикл развития Fucus vesiculosus
Фукус зубчатый (Fucus serratus) имеет ветви с зубчатыми краями. Талломы до полуметра или метра высотой, бурые, сильно разветвленные. Ветви широкие, в верхней части вздувшиеся, без типичных воздушных пузырей. Рецептакулы плоские, не отграниченные от ветвей с зубчатым краем, иногда с вильчатыми верхушками. Растет на границе литорали и сублиторали на каменистых грунтах. Встречается у берегов Европы и Северной Америки, в морях России – Белом и Баренцевом.
У водорослей рода саргассум (Sargassum) (рис. 109: 3) слоевище кустистое с обильными ветвящимися веточками. От ветвей берут свое начало листовидные пластинки с четко выраженным срединным ребром или без него. Воздушные пузыри имеются на веточках, развивающихся в пазухах листовидных пластинок. В тропиках есть однолетние виды. Растение прикрепляется к грунту конической подошвой. Представители рода распространены в основном в тропических и субтропичесикх морях и в теплых водах морей умеренных широт, в России – в дальневосточных морях. В Саргассовом море обитают виды, лишенные органов прикрепления, образующие огромные массы, плавающие в поверхностных слоях воды. В оогонии одна яйцеклетка.
205
Аскофиллум узловатый (Ascophyllum nodosum) (рис. 109: 4) имеет раздельнополые слоевища до 1,5 м длиной сильно дихотомически или неправильно разветвленные. На ветвях попеременно или супротивно расположены сдавленно-цилиндрические мясистые более короткие ветви без средней жилки. По длине ветви имеют расширения – воздушные пузыри, располагающиеся поодиночке. Рецептакулы развиваются на коротких веточках по бокам длинных ветвей и при созревании становятся похожими на крупные оранжевые ягоды. Таллом многолетний; растение произрастает на каменистых и песчаных грунтах в защищенных от прибоя местах в литоральной зоне в Белом и Баренцевом морях.
Следующие отделы содержат в своих чисто зеленых хлоропластах хлорофиллы a и b, пластиды приобретены за счет первичного и вторичного эндосимбиоза.
йЪ‰ВО Chlorophyta – бВОВМ˚В ‚У‰УрУТОЛ
ä этому отделу относятся около семи тысяч видов микроскопических и макроскопических эукариотических водорослей, окраска которых напоминает окраску высших растений. Отдел вклю-
чает одноклеточные, колониальные и многоклеточные формы всех известных у водорослей типов морфологической организации таллома, кроме амебоидной. Пигментная система хлоропластов образована хлорофиллами а и b, преобладающими над каротиноидами (α-, β-, γ-, ε- каротинами и ксантофиллами: лютеином, неоксантином, виолаксантином, зеаксантином, антераксантином). Хлорофиллы а и b имеются еще у прохлорофитовых (прокариоты), а среди эукариот – у хлорарахниофи-
товых (отдел Chlorarachniophyta), стрептофитовых (отдел Streptophyta)
и эвгленовых (отдел Euglenophyta).
Каротиноиды не маскируют зеленую окраску хлорофилла, за счет чего хлоропласты окрашены в чисто зеленый цвет. Хлоропласты покрыты двумя мембранами, как у глаукофитовых, красных, стрептофитовых водорослей и высших растений. Тилакоиды сгруппированы в граны, опоясывающей ламеллы нет. Пиреноиды, если имеются, расположены внутри хлоропласта. Запасной продукт – углевод крахмал – откладывается внутри хлоропласта: вокруг пиреноида и в строме. У дазикладовых
206
водорослей (Dasicladaceae) вместо крахмала запасается инулин и другие углеводы, у части видов – масла, редко глицерин. Клеточная стенка состоит преимущественно из целлюлозы, реже встречается маннан, ксилан и др. У ряда форм в оболочках клеток найден спорополленин, характерный для высших растений, у которых он входит в состав покрова спор и пыльцы. У некоторых представителей зеленых водорослей, например, эдогониума (Oedogonium) на поверхности клеточной оболочки выделяется кутин, образующий твердую кутикулу. У части видов оболочка пропитывается солями железа (например, у факотуса – Phacotus) или кальция (некоторые сифоновые).
Клеточная оболочка цельная или состоит из отдельных фрагментов, например у некоторых вольвоксовых. Очень часто оболочка инкрустирована шипиками, гранулами, бородавками, щетинками и другими украшениями. В оболочке образуются поры, через которые протопласт осуществляет связь с внешней средой. Через поры выделяется слизь, способствующая облегчению остаточной массы клетки у планктонных видов, способствующая образованию колоний, активному передвижению клетки в пространстве. Через поры протопласты у некоторых колониальных и многих многоклеточных видов сообщаются между собой посредством плазмодесм. Нередко слизь покрывает всю клетку в связи с ослизнением внешних слоев оболочки, иногда наблюдаются слизистые капсулы. При неблагоприятных условиях отмечается временное ослизнение клеточных стенок и переход водорослей в пальмеллевидное состояние.
В процессе вегетативного деления клеток образование поперечных перегородок осуществляется преимущественно с участием фикопласта. Фикопласт – система микротрубочек, которые во время телофазы собираются в экваториальной плоскости между разделившимися дочерними ядрами (рис. 25: 1, 2). Клетки зеленых водорослей содержат от одного до многих ядер. Ядра с одним, несколькими или многими ядрышками.
У монадных форм имеется стигма, являющаяся частью хлоропласта. Она состоит из одного или нескольких слоев пигментных глобул, расположенных у поверхности хлоропласта под его оболочкой. Функцию фоторецепции, по-видимому, выполняет участок наружной мембраны хлоропластной оболочки или плазмалеммы, прилегающий к стигме. Жгутики у монадных форм и стадий развития обычно в числе 2–4 изоконтные (одинаковые по длине), изодинамичные, гладкие (у некоторых видов жгутики покрыты тонкими волосками или мелкими чешуйками). Иногда жгутик может быть один или их число возрастает до многих
207
(монадные стадии Oedogonium, зооспоры Derbesia). У празинофитовых жгутики выходят из особой жгутиковой ямки, находящейся на переднем конце клетки, или из субапикального желобкообразного углубления.
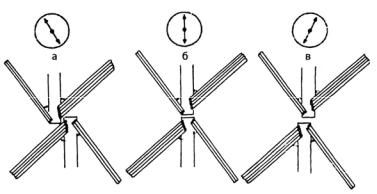
Жгутики в клетке чаще всего прикрепляются по крестообразному типу. При этом четыре крестовидно расположенных микротрубочковых корня закрепляют базальные тела жгутиков в клетке. Если смотреть на клетку со жгутиками сверху, два базальных тела могут располагаться в пространстве как цифры 11 и 5 на циферблате часов (тип расположения 11–5). У водорослей этого отдела встречается также расположение базальных тел жгутиков по типу 12–6 и 1–7 (рис. 110).
Рис. 110. Расположение базальных тел и жгутиковых корней у Chlorophyta: а – тип 11–5, базальные тела слегка сдвинуты относительно друг друга против часовой стрелки; б – тип 12–6, гипотетический исходный жгутиковый аппарат, у которого базальные тела расположены на одной линии; в – тип 1–7, базальные тела слегка сдвинуты относительно друг друга по часовой стрелке
Размножение вегетативное, бесполое и половое. У одноклеточных, лишенных клеточной оболочки (например, дуналиеллы – Dunaliella), размножение осуществляется простым делением клетки надвое. У колониальных и многоклеточных вегетативное размножение осуществляется фрагментацией колоний или талломов. Ценобиальные зеленые водоросли (вольвокс – Volvox, гидродикцион – Hydrodictyon) образуют дочерние ценобии внутри материнских. У многих зеленых водорослей отмечается образование акинет.
208
Бесполое размножение осуществляется спорами различного строения и в жизненном цикле зеленых водорослей присутствует, наряду с вегетативным и половым размножением. В основном выделяют подвижные зооспоры и неподвижные апланоспоры. И те, и другие образуются внутри спорангиев, при этом оболочка спор формируются без участия оболочки спорангия. Если апланоспоры внутри спорангия приобретают форму материнской клетки, то они называются автоспорами. Апланоспоры с утолщенной оболочкой, способные длительное время быть в состоянии покоя, называются гипноспорами. Гемизооспоры – это зооспоры, утрачивающие жгутики, но сохранившие сократительные вакуоли и стигму.
Половой процесс разнообразен: хологамия (например, у дуналиеллы
–Dunaliella), изогамия (некоторые виды рода хламидомонас – Chlamydomonas), гетерогамия или анизогамия (некоторые виды рода эвдорина
–Eudorina), оогамия (вольвокс – Volvox). Зиготы, образующиеся при холо-, изо- и гетерогамии, обычно проходят стадию планозиготы (подвижной зиготы) и гипнозиготы (покоящейся зиготы). При оогамии образуются неподвижные зиготы, которые обычно проходят стадию гипнозиготы (ооспоры, зигоспоры) (например, у эдогониума – Oedogonium). Иногда зигота без стадии покоя сразу прорастает в новое растение (многие Bryopsidales). Среди представителей зеленых водорослей имеются все типы циклов развития (гаплонты, диплонты, изо- и гетероморфная смена форм развития).
Общепринятой системы зеленых водорослей нет, поскольку до сих пор отсутствуют объективные критерии (морфологические, ультраструктурные, биохимические, генетические) выделения крупных таксонов, начиная с классов и выше. Деление зеленых водорослей на классы в разных системах сильно различается, и их общее количество доходит до 8–9, особенно в последние годы. В данном пособии принята система зеленых водорослей, приведенная в учебнике П. Зитте с соавторами (2007). Отдел зеленые водоросли – Chlorophyta включает классы: прази-
нофициевые – Prasinophyceae, ульвофициевые – Ulvophyceae, тре-
буксиофициевые – Trebouxiophyceae, кладофорофициевые – Cladophorophyceae, бриопсидофициевые – Bryopsidophyceae, дазикладофициевые
–Dasycladophyceae, трентеполиофициевые – Trentepohliophyceae, хло-
рофициевые или зеленые водоросли – Chlorophyceae.
209
К классу Prasinophyceae – Пра-
зинофициевые водоросли – отно-
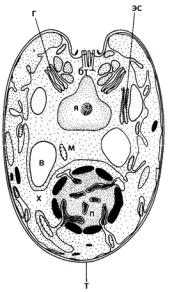
сятся монадные, пальмеллоидные (капсальные) и коккоидные формы, одиночные или образующие колонии. У монадных клеток их поверхность и жгутики покрыты одним или несколькими слоями чешуек до пяти различных по морфологии типов. Предполагается, что эволюционная сложность клетки возрастает по мере увеличения числа слоев чешуек. У более высокоорганизованных водорослей, например, тетрасельмиса (Tetraselmis) (рис. 111), эти слои объеди-
нены между собой в жесткий покров типа теки или оболочки.
Одноядерные клетки содержат крупный лопастной пристенный хлоропласт, как правило, с пиреноидом. В состав пигментной системы празиновых входит специфический ксантофилл – сифоноксантин, редко встречающийся у других зеленых водорослей (например, у бриопсидовых). Основными продуктами запаса являются крахмал и маннитол (органическое соединение из класса спиртов). У многих представителей име-
ется стигма, у некоторых – стрекательные органеллы – трихоцисты. Жгутиков разное количество, чаще – 2–4. У одножгутиковых и двужгутиковых форм они прикрепляются сбоку, у четырехжгутиковых – ундулиподии примерно одинаковой длины и выходят из особого углубления в апикальной части сердцевидной клетки. Некоторые виды имеют чередующиеся подвижную и неподвижную стадии. Бесполое размножение происходит с образованием зооспор или путем простого деления клетки надвое; половое размножение известно только для одного вида.
Большая часть представителей этого класса являются свободноживущими обитателями солоноватых и пресных водоемов. Некоторые ви-
210