

Основы альгологии и микологии Охапкин
.pdfобычно заметные. Акинеты всегда отсутствуют. Виды рода встречаются в стоячих и текучих водах, на орошаемых скалах, в зоне прибоя озер, на водных растениях, на камнях, раковинах моллюсков, а также в болотах и солоноватых водоемах, реже – на сырой почве, некоторые виды предпочитают воды, богатые известью.
У представителей порядка Stigonematales – Стигонемовые в ре-
зультате поперечных и продольных клеточных делений формируются однорядные или многорядные нити, характерно настоящее ветвление. Могут встречаться гетероцисты и акинеты.
Цианопрокариоты встречаются большинстве биотопов и распространены по всему земному шару. Обитают в морских и пресных водоемах как компоненты планктона, бентоса и обрастаний различных субстратов. Есть виды, развивающиеся на поверхности снега, влажных скал, почвы, формирующие симбиозы с другими организмами. Принимают участие в образовании известковых пород. Ископаемые слоистые известковые отложения (строматолиты) известны из докембрийских отложений, и полагают, что образовавшие их цианопрокариоты уже в ту геологическую эпоху были широко распространены и сами отложения занимали обширные площади.
Многие виды синезеленых водорослей вызывают «цветение» воды и являются токсичными для животных и человека. Огромна роль цианопрокариот в фиксации атмосферного азота и в переводе этого важнейшего для функционирования живых организмов элемента в форму, удобную для их жизнедеятельности.
йЪ‰ВО Prochlorophyta – ирУН‡рЛУЪЛ˜ВТНЛВ (ФВр‚Л˜М˚В) БВОВМ˚В ‚У‰УрУТОЛ, ЛОЛ ирУıОУрУЩЛЪУ‚˚В ‚У‰УрУТОЛ
ùтот отдел прокатиотических фотосинтезирующих с выделением кислорода организмов выделен в 1976 г. Р. Левиным. Пред-
ставители рода прохлорон (Prochloron) живут в симбиозе с разными видами асцидий, обитающих в тропических и субтропических морских литоральных водах. Особи Prochloron обитают на поверхности и внутри колоний асцидий, в их клоаке или в материале туники. Животные от
91
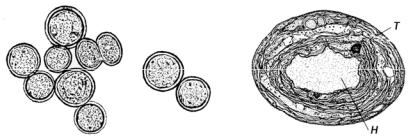
присутствия клеток водорослей принимают зеленый цвет. Вне асцидий эти водоросли обычно не встречаются.
Индивиды прохлорон одноклеточные, часто образуют группы, но без слизистых обверток вокруг клеток или колониальной слизи (рис. 50).
Рис. 50. Внешний вид индивидов Prochloron и строение клетки по данным электронной микроскопии: Т – тилакоид; Н – нуклеоид
Клетки окрашены в ярко-зеленый цвет за счет пигментов хлорофиллов а и b, найдены также пигмент, подобный хлорофиллу с, и каротиноиды. Фикобилиновые пигменты, характерные для синезеленых водорослей, отсутствуют. Размножаются делением клетки надвое. Клетки шаровидные, после деления полушаровидные, 6-30 мкм в диаметре. Клеточная стенка многослойная, содержит муреин. Ядер и других клеточных органелл, окруженных мембранами, нет. ДНК находится в нуклеоплазматическом районе (нуклеоиде). Размер генома такой же, что и у синезеленых водорослей. Обнаружены тилакоиды, расположенные парами.
Прохлорококкус (Prochlorococcus) известен как один из основных компонентов планктонного комплекса поверхностных слоев Атлантического, Индийского, Тихого океанов, найден в Саргассовом и Средиземном морях, а также в Персидском заливе. Клетки имеют эллипсоидную форму и по размеру (диаметр 0,5–0,6 мкм и длина 0,7–0,8 мкм) намного меньше большинства хроококковых синезеленых водорослей. Центральную область клетки занимает нуклеоид, содержащий ДНК, на периферии параллельными рядами лежат тилакоиды. Отличается от других представителей этой группы особым набором хлорофиллоподобных пигментов.
Прохлоротрикс (Prochlorothrix) – это свободноживущий пресноводный организм, выделенный из озера в окрестностях г. Амстердам и
92
внешне сходный с цианеями порядка осцилляториевых. Особи имеют вид коротких нитей, состоящих из цилиндрических клеток длиной 3–10 мкм и диаметром 0,5–1,5 мкм.
Относительно сходства прохлорофитовых с другими организмами единого мнения нет. Прокариотические зеленые водоросли по строению клетки близки к бактериям и с современных позиций должны рассматриваться как бактерии. Морфологические признаки и способность к оксигенному фотосинтезу сближает их с синезелеными водорослями. Молекулярно-биологические данные последних лет свидетельствуют об их близком родстве с некоторыми цианопрокариотами, и в современных системах они рассматриваются среди синезеленых. При этом однокле-
точные формы помещают в Chroococcales, нитчатые – в Oscillatoriales.
Промежуточные черты строения прохлорофитовых водорослей и их физиолого-биохимические особенности дали основание для различных гипотез, касающихся их происхождения и вероятных путей эволюции. Например, допускают, что они произошли от синезеленых водорослей путем утраты фикобилинов и приобретения хлорофилла b. Другие исследователи считают, что становление Prochlorophyta шло параллельно со становлением отдела Cyanophyta, и выводят прохлорофитовые от фотосинтезирующих бактерий. Одни авторы рассматривают прохлорофитовые как промежуточное звено на пути развития эукариотических водорослей из синезеленых. Другие, придерживаясь теории симбиогенеза, считают древние прохлорофитовые водоросли вероятными предками хлоропластов высших растений, которые могли возникнуть в результате внедрения прокариотических зеленых водорослей в клетки неспособного к фотосинтезу гетеротрофного хозяина. Однако сравнение последовательностей 16S рибосомальной РНК не позволяет сближать прохлорон с зелеными водорослями и высшими растениями. Некоторые ученые считают, что хлорофиллы а и b могли возникать в процессе эволюции неоднократно, и поэтому не обязательно считать прохлорофитовых предками зеленых водорослей и высших растений. В связи с этим, прохлорофитовые водоросли являются наиболее интересными с точки зрения решения проблем эволюционного происхождения эукариотической растительной клетки.
93
éÚ‰ÂÎ Glaucophyta –
ЙО‡ЫНУЩЛЪУ‚˚В ‚У‰УрУТОЛ (ЙО‡ЫНУЩЛЪ˚)
Éмического положения, включающая немногих представителей коккоидной и монадной структуры, у которых хлоропластылаукофитовые водоросли – сборная группа неясного таксоно-
трактовались как цианопрокариоты, ведущие эндосимбиотический образ жизни и получившие название цианеллы. Хлоропласты глаукофитовых содержат хлорофилл а и фикобилиновые пигменты (как у синезеленых), хлорофилл b отсутствует. Фикобилиновые пигменты находятся в фикобилисомах, расположенных внутри хлоропластов на тилакоидах, не сгруппированных между собой. В клетках глаукофитовых водорослей хлоропласты располагаются либо упорядоченно в виде двух групп, либо беспорядочно, случайно. Включения запасных питательных веществ, представленные у синезеленых различными метаболическими гранулами, не наблюдаются. Сходство хлоропластов глаукофитовых со свободно живущими цианопрокариотами проявляется в наличии тонкой оболочки, содержащей муреин. Однако размер их генома составляет одну десятую генома цианобактерий и соответствует размеру генома хлоропластов других эукариотных водорослей. Строение генома и потеря способности к размножению и культивированию вне клетки хозяина свидетельствует в пользу трактовки цианелл глаукофитовых как истинных хлоропластов с чертами примитивной организации. Структура жгутиков у монадных представителей имеет общие черты со жгутиками зеленых водорослей.
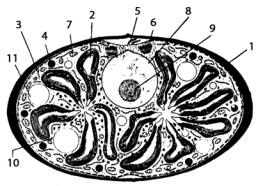
С позиций ультраструктурной организации клетки хорошо изучен глаукоцистис (Glaucocystis nostochinearum) (рис. 51).
Рис. 51. Схема строения клетки Glaucocystis:
1 – клеточная стенка;
2 – хлоропласт;
3 – эндоплазматический ретикулум; 4 – липидная глобула;
5 – рудиментарный жгутик; 6 – диктиосома; 7 – митохондрия; 8 – ядро; 9 – ядрышко;
10 – крахмал; 11 – вакуоль
94
Это типично эукариотическая водоросль, в которой находятся коккоидные цианобактерии, сходные по структуре с цианопрокариотой Aphanothece, рассматриваемые как хлоропласты. Клетки имеют толстую клеточную стенку, ядро и другие органеллы, типичные для эукариотических водорослей. У клеток бесцветного организма – хозяина глаукоцистиса – утрачивается способность к самостоятельному образованию крахмала. Оболочка хлоропластов глаукоцистиса сильно редуцирована, но в ее составе обнаруживается остаточный пептидогликановый слой. В отличие от настоящих хлоропластов, присущих остальным эукариотическим водорослям, распределение хлоропластов между дочерними клетками глаукоцистиса в процессе деления материнской клетки носит случайный характер и поэтому их число в них сильно варьирует.
Глаукофитовые – это живые примеры возможного строения предковых жгутиковых форм водорослей; их существование в природе и особенности строения выдвигаются сторонниками эндосимбиотической теории происхождения хлоропластов в качестве весомых аргументов в ее пользу.
йЪ‰ВО Rhodophyta – др‡ТМ˚В ‚У‰УрУТОЛ
äрасные водоросли, или багрянки, – преимущественно морские растения. Это самая обширная группа растений – около четы-
рех тысяч видов, относящихся более чем к шестистам родам. В пресных и солоноватых водах, а также во вневодных местообитаниях обитает немногим более пятидесяти видов. Водоросли прикрепленные, преимущественно многоклеточные, макроскопические нитчатой, разнонитчатой, паренхиматозной (пластинчатой) или ложнопаренхиматозной структуры. Реже водоросли одноклеточные или колониальные, коккоидной структуры, микроскопических размеров. Подвижные стадии в жизненном цикле отсутствуют.
Окраска водорослей от розовой, красной и оранжевой до оливковозеленой, сине-зеленой, фиолетовой, почти черной. Есть бесцветные паразитические виды. Пластиды красных водорослей по строению напо-
95
минают пластиды глаукофитовых. Окраска обусловлена наличием в хлоропластах помимо хлорофилла а (хлорофиллов b и c нет, наличие хлорофилла d долгое время считалось спорным), каротиноидов (α- и β- каротина, зеаксантина, лютеина, антераксантина, криптоксантина, неоксантина) и фикобилинов – красного фикоэритрина и синих фикоцианина и аллофикоцианина. Фикобилиновые пигменты – это дополнительные пигменты, позволяющие фотосинтезировать при слабой интенсивности освещения, особенно в глубоко проникающих в воду синих лучах. У морских видов все пигменты большей частью маскируются фикоэритрином. У пресноводных багрянок фикоэритрина мало, поэтому они обычно окрашены в серый, темно-серый, темно-зеленый, сине-зеленый или ярко-зеленый цвет. Цвет талломов зависит от глубины обитания водорослей. Например, порфира (Porphyra, рис. 52: 3), растущая на незначительной глубине, имеет зеленовато-красноватый оттенок, а с погружением на большие глубины становится интенсивно багряной. Объяснение этому явлению дал немецкий ученый Т.В. Энгельман. Поскольку наибольшей проникающей способностью обладают лучи синефиолетовой части спектра, достигающие самых больших глубин, то у глубоководных багрянок в составе пигментов увеличивается процент фикоэритрина, и таким образом водоросли более успешно поглощают сине-фиолетовую часть спектра. Багрянки – это наиболее глубоководные фотосинтезирующие организмы. На каменистых плитах в районе Багамских островов были обнаружены заросли красных водорослей на глубине 268 м. На эту глубину проникает только 0,0005% солнечной радиации, падающей на поверхность воды.
Слоевище красных водорослей нитевидное, кустистое, пленчатое, корковидное или пластинчатое, шнуровидное, трубчатое или иной фор-
мы (рис. 52).
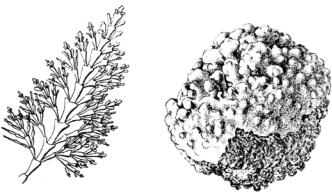
Нитевидные талломы могут быть очень тонкими, волосовидными, состоящими из однорядных нитей, либо более грубыми, состоящими из нескольких рядов клеток. Цилиндрические талломы плотные или с полостью внутри, слизистые, хрящеватые или кожистые, в виде шнуров или обильно ветвящихся кустиков. Пластинчатые талломы могут быть цельными, сложно рассеченными с дополнительными выростами по краю и поверхности таллома. Тонкие, шнуровидные талломы наблюдаются, например, у немалиона (Nemalion, рис. 52: 1), талломы в виде ветвящихся кустиков – у каллитамниона (Callithamnion, рис. 52: 2), пластинок разной величины – порфиры (Porphyra, рис. 52: 3).
96

Рис. 52. Форма слоевищ красных водорослей:
1 – Nemalion; 2 – Callithamnion; 3 – Porphyra; 4 – Delesseria; 5 – Phycodris; 6 – Batrachospermum; 7 – Polysiphonia
У некоторых красных водорослей талломы сложно рассечены и имеют как бы жилки, и эти водоросли внешне напоминают листья высших растений, например, делессерия (Delesseria, рис. 52: 4) или фикодрис (Phycodris, рис. 52: 5). Водоросли, имеющие вид корок или пленок, часто развиваются как эпифиты на других водорослях. Пресноводные формы мелкие, нередко микроскопические, одноклеточные, колониальные или нитчатые, не достигают такого морфологического разнообразия, как морские.
В основе нитчатого одноосевого таллома у красных водорослей лежит однорядная осевая нить с боковыми веточками ограниченного роста (например, батрахоспермум – Batrachospermum, рис. 52: 6; 59). При так называемом многоосевом (фонтанном) строении в центре слоевища находится пучок параллельных осевых нитей, окруженных снаружи боковыми веточками (полисифония – Polysiphonia, рис. 52: 7; 62). Часть
97
видов прикрепляется к субстрату ризоидами или подошвой. Нарастание таллома в основном верхушечное.
Форма клеток красных водорослей – шаровидная, яйцевидная, боченкообразная, цилиндрическая или многоугольная. Оболочка клетки цельная двухслойная. Внутренний слой состоит из целлюлозных волокон. Оболочка содержит много пектиновых веществ и гемицеллюлоз, поэтому сильно набухает в воде и частично ослизняется. Клеточные оболочки часто содержат так называемые фикоколлоиды – агар, агароиды, каррагенин. Это сложная смесь полисахаридов, содержащих серу и сильно набухающих в воде. В горячей воде эти вещества могут растворяться, а при охлаждении растворов дают студни. Эти вещества служат объектом промысла, в основном добывают агар. У некоторых красных водорослей кнаружи от оболочки откладывается кутикула. На мягких частях слоевища или на мягких слоевищах кутикулы нет, наружная часть оболочки желатинизированная или ослизненная. Иногда клеточные оболочки могут инкрустироваться известью; у водорослей семейства Corallinaceae откладывается кальцит (карбонат кальция с примесью карбонатов магния, железа и марганца), у некоторых Nemaliales – арагонит – минерал класса карбонатов. Слоевище таких водорослей приобретает консистенцию камня, например у кораллины (Corallina) и лито-
тамниона (Lithothamnion) (рис. 53).
Рис. 53. Corallina (слева) и Lithothamnion
У Corallina officinalis обызвествленный таллом сохраняет форму кустика. Обызвествление захватывает поверхностные коровые части члеников, а сочленения не обызвествлены и таллом сохраняет подвиж-
98
ность, веточки могут изгибаться под действием течений. У Lithothamnion формируются либо кустики, либо скопления, подобные камню или кораллам, формируются и корковидные, и кустовидные талломы. Виды этих родов принимают участие в формировании коралловых рифов в тропических морях.
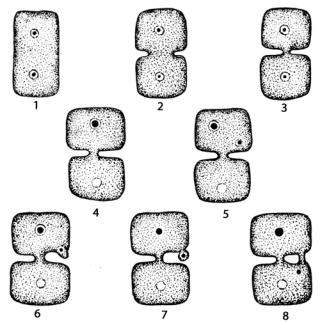
Клетки в талломе красных водорослей соединены с помощью первичных и вторичных пор (рис. 54).
Рис. 54. Схема образования первичных (1–3) и вторичных (4–8) пор у красных водорослей
При делении клеток у большинства красных водорослей (все бангиевые и примитивные представители класса флоридеевые) в результате неполного смыкания растущей от периферии к центру клеточной стенки образуются первичные поры. Затем эта пора закрывается пробкой из белков и полисахаридов. Через первичные поры проходят тяжи, соединяющие цитоплазму соседних клеток. Кроме первичных, у большинства
99
флоридей происходит вторичное соединение соседних клеток одной или несколькими вторичными порами.
Клетки красных водорослей содержат все органеллы, присущие эукариотам. Хлоропласты у более примитивных представителей одиночные звездчатой формы с пиреноидом. У флоридеевых водорослей хлоропласты лентовидной, зернистой, линзовидной, дисковидной, чашевидной и другой формы, как правило, без пиреноидов. Хлоропласт окружен двумя оболочками, содержит одиночные тилакоиды, ориентированные обычно параллельно друг другу. На поверхности тилакоидов локализованы дисковидные или шаровидные тельца – фикобилисомы. Периферических опоясывающих ламелл нет у примитивных форм со звездчатыми или лопастными хроматофорами, ноониесть увсех флоридей.
Основной продукт ассимиляции – багрянковый крахмал – откладывается в цитоплазме или на поверхности хлоропластов вне связи с пиреноидами. От йода он приобретает буро-красный цвет. По химической природе он ближе к амилопектину и гликогену, при гидролизе дает галактозу. Реже откладываются сахариды и многоатомные спирты.
Вегетативное размножение красных водорослей осуществляется фрагментацией таллома, у части видов наблюдается образование дополнительных, отрастающих от подошвы или стелющихся по субстрату побегов. Бесполое размножение происходит посредством моно-, би-, тетраили полиспор – неподвижных клеток, развивающихся по одной, две, четыре или много в спорангиях. Моноспоры развиваются у Bangiophyceae в клетках, называемых моноспорангиями, в результате превращения всего протопласта клетки в спору, лишенную оболочки, с одним ядром и одним хлоропластом. У флоридей моноспоры образуются только в порядке Nemaliales. Моноспоры образуются в любой клетке таллома (у бангиевых) или на веточкахограниченногороста(участифлоридей).
У большинства флоридеевых водорослей наиболее обычны тетраспоры¸ образующиеся в тетраспорангиях. Тетраспоры образуются на диплоидных бесполых растениях – тетраспорофитах. При образовании тетраспор происходит мейоз. Расположение тетраспор в тетраспорангиях может быть крестообразным (гелидиум – Gelidium, хондрус – Chondrus), рядовым (кораллина – Corallina) или по углам тетраэдра (церамиум – Ceramium). Тетраспорангии формируются из конечных клеток веточек ограниченного роста и выглядят как боковые выросты вертикальных нитей. Часто тетраспорангии в процессе роста оказываются погруженными в коровый слой. У некоторых видов тетраспорангии развиваются в нематециях, имеющих вид небольших подушечек на поверхно-
100