

Основы альгологии и микологии Охапкин
.pdfРис. 22. Диаграмма классификации митозов
Полузакрытый митоз характеризуется образованием в ядерной оболочке полярных отверстий или фрагментацией ядерной оболочки, но полностью она не исчезает. Открытый митоз – ядерная оболочка полностью исчезает, как у высших растений. При митозе варьирует и форма веретена деления. У некоторых водорослей ядрышко в период митоза не исчезает, а только разрыхляется, распределяясь по ядру (например, у спирогиры – Spirogyra), у других (например, у эвгленовых) делится как самостоятельная структура. У цианопрокариот и прохлорофитовых морфологически оформленное ядро отсутствует. Нити ДНК расположены в центральной части клетки – нуклеоиде, не связаны с гистонами в структуры, подобные хромосомам эукариотических водорослей.
51
Динофитовые водоросли имеют заметные отличия в строении и химическом составе хромосом в сравнении с другими эукариотами (например, в хромосомах гистоны либо не обнаруживаются, либо их содержание крайне низко). В интерфазных ядрах хромосомы динофитовых остаются спирализованными и хорошо заметны. Имеются также отличия в ходе митоза и цитокинеза.
Таким образом, в цитоплазме эукариотических водорослей формируется единая система взаимосвязанных и взаимопревращающихся мембранных структур, к которым относятся мембраны ядра, пластид, митохондрий, аппарата Гольджи, эндоплазматической сети и производных от них – вакуолей и микротел. Они делят цитоплазму на самостоятельно функционирующие специализированные отсеки – органеллы. Клетки прокариотических оксигенных фотосинтетиков (цианопрокариот и прохлорофитовых) не имеют такой взаимной системы мембранных структур. Их клетки богаты включениями, среди которых следует назвать зерна гликогеноподобного полисахарида, кристаллы цианофицина – специфического белка синезеленых, волютина – запасы полифосфатов. Разнообразные по составу включения имеются также и в клетках эукариотических водорослей. Так, например, зеленые водоросли запасают крахмал, который накапливается в пластидах. Эвгленовые вырабатывают особый полисахарид – парамилон, концентрирующийся вокруг пиреноида или непосредственно в цитоплазме. У золотистых, желтозеленых, реже диатомовых водорослей в цитоплазме накапливается хризоламинарин, который в мертвых клетках разрушается. Для бурых водорослей характерен водорастворимый ламинарин, для красных – багрянковый крахмал. У многих водорослей откладываются глобулы липидов, особенно у диатомовых и динофитовых. У многих групп в цитоплазме присутствуют плотные сферические структуры, содержащие полифосфаты – волютиновые зерна.
Деление клетки – важнейший процесс, лежащий в основе размножения у одноклеточных водорослей и роста многоклеточных. Он тесно связан с митозом. В зависимости от степени согласованности митоза и цитокинеза различают последовательное (сукцессивное) деление, при котором за делением ядра сразу же следует образование клеточной оболочки (рис. 23).
52
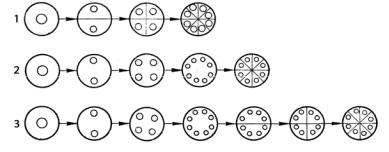
Рис. 23. Способы клеточного деления у водорослей:
1 – последовательное (сукцессивное); 2 – одновременное (симультанное); 3 – постепенное (сегрегативное)
При одновременном (симультанном) делении образование клеточных перегородок происходит одновременно после завершения всей серии ядерных делений в клетке. Есть еще постепенное (сегрегативное) деление, которое осуществляется в несколько этапов. Сначала ядро клетки многократно делится и образуется многоядерный или ценоцитный таллом, который способен к росту и дифференцировке. Затем по всему таллому или только на его отдельных участках формируются перегородки, делящие его на многоядерные сегменты, которые в свою очередь подвергаются делению. В результате этих процессов формируется таллом, состоящий из одноядерных или многоядерных клеток или сегментов. Такой тип деления лежит в основе формирования сифонокладальной структуры таллома, наблюдающейся, например, у зеленых водорослей из порядка Cladophorales (Siphonocladales). Считают, что разобщение во времени процессов митоза и цитокинеза могло стать причиной появления у водорослей сифональной структуры таллома.
Клеточное деление эукариотных организмов – это процесс, при котором одна клетка делится на две или несколько частей (дочерних клеток) в результате митоза и цитокинеза. Простейшим случаем клеточного деления является деление надвое путем перешнуровки голых, лишенных оболочки одноклеточных жгутиковых водорослей, например дуналиеллы (Dunaliella), пединомонаса (Pedinomonas) или одноклеточных жгутиковых с клеточным покровом (эвгленовые, динофитовые, криптофитовые) (рис. 24). Этот процесс называется схизотомией. Схизотомия обеспечивает равномерное распределение между
53
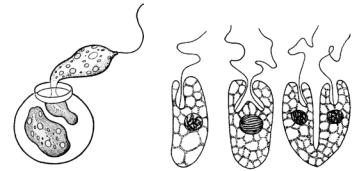
двумя дочерними организмами всех компонентов материнской клетки, включая клеточные покровы, например, пелликулы у эвгленовых или теки у динофитовых. Схизотомия лежит в основе бесполого размножения одноклеточных безоболочковых фитомонад или процесса образования колоний.
Рис. 24. Деление клетки
уэвгленовых водорослей Trachelomonas (слева)
иRhabdomonas (справа)
Собразованием у водорослей клеточной оболочки они утрачивают способность к схизотомии. При этом цитокинез водорослей может происходить без участия оболочки материнской клетки, например, при формировании зооспор, или гамет у вольвоксовых. При образовании автоспор многих водорослей их оболочки формируются независимо от оболочки материнской клетки. Этот процесс называется споруляцией, или элеутеросхизисом, или схизогонией. У других водорослей в формировании оболочки дочерних клеток принимают участие только внутренние слои оболочки материнской клетки. Когда вся оболочка материнской клетки принимает участие в формировании оболочек дочерних клеток, такой процесс называется десмосхизисом. Десмосхизис происходит при делении вегетативных клеток надвое в многоклеточном талломе. Десмосхизис протекает в трех взаимно перпендикулярных плоскостях. В отличие от споруляции, десмосхизис не ведет к образованию спор и бесполому размножению с помощью специализированных клеток, а к росту слоевища. Этот процесс может впоследствии сопровождаться вегетативным размножением путем фрагментации таллома. Не-
54
редко десмосхизис, обеспечивающий рост слоевища, сопровождается споруляцией, ведущей к бесполому размножению.
Унитчатых форм клеточное деление происходит по типу цитотомии.
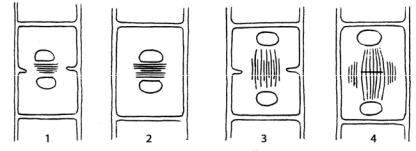
Уводорослей цитотомия может происходить с помощью кольцевой борозды деления, развивающейся от периферии к центру или путем формирования клеточной пластинки, которая растет от центра к периферии клетки (рис. 25). Эти типы разделения могут сочетаться с образованием так называемого фрагмопласта – системы микротрубочек, ориентированных параллельно оси деления клетки, и фикопласта – системы микротрубочек, расположенных перпендикулярно оси веретена и параллельно плоскости деления клетки. Вариантов цитотомии у водорослей много, на чем некоторые альгологи основывают деление их на крупные таксономические группы.
Рис. 25. Типы цитокинеза у зеленых водорослей (в широком смысле): 1, 2 – интерзональное веретено отсутствует, ядра тесно сближены; 3, 4 – интерзональное веретено сохраняется, ядра удалены друг от друга
(1 – образование кольцевой борозды деления и фикопласта; 2 – образование клеточной пластинки с участием фикопласта; 3 – образование кольцевой борозды деления; 4 – образование клеточной пластинки
с участием фрагмопластоподобной структуры)
Важным понятием для водорослей является понятие вегетативного клеточного деления. Это такое деление одетых оболочками клеток водорослей, которое приводит к образованию вегетативных клеток, а не репродуктивных – спор, и способствует росту слоевища. Таким образом, десмосхизис и цитотомия – это две разновидности вегетативного клеточного деления, причем десмосхизис, по-видимому, возник раньше цитотомии.
55
нЛФ˚ ПУрЩУОУ„Л˜ВТНУИ Ур„‡МЛБ‡ˆЛЛ Ъ‡ООУП‡ ‚У‰УрУТОВИ
Ç одоросли – это одноклеточные, колониальные или многоклеточные организмы. Тело водорослей не расчленено на органы и представлено талломом или слоевищем. Слоевище водорослей
различных отделов устроено разнообразно, поэтому выделяют несколько основных типов морфологической дифференциации талломов.
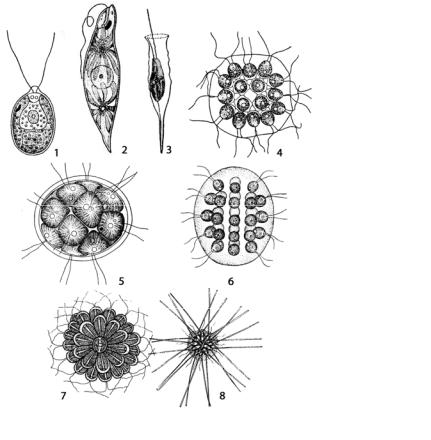
Монадный (жгутиковый) тип морфологической дифференциации талломахарактеризуетсяактивнойподвижностьюспомощьюжгутиков(рис. 26).
Рис. 26. Монадный (жгутиковый) тип морфологической дифференциации таллома у водорослей: 1 – Chlamydomonas;
2 – Euglena;
3 – Dinobryon;
4 – Gonium;
5 – Pandorina,
6 – Eudorina;
7 – Synura;
8 – Chrysosphaerella
56
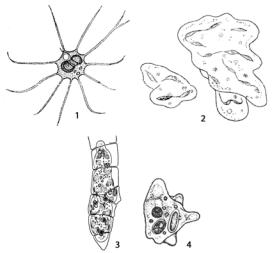
Подвижность характерна для одноклеточных жгутиковых водорослей, встречающихся во многих отделах (например, у зеленых вольвок-
совых – Chlamydomonas, эвгленовых – Euglena и других) (рис. 26: 1, 2).
Разновидность монадной организации – подвижные (с помощью жгутиков) колонии, состоящие из неопределенного числа клеток (например,
золотистые водоросли Dinobryon, Synura, Chrysosphaerella) (рис. 26: 3, 7, 8), или ценобии – колонии, имеющие определенное (обычно кратное двум) число клеток. Ценобии наблюдаются у зеленых водорослей
Gonium, Pandorina, Eudorina (рис. 26: 4–6) и других. У более высоко ор-
ганизованных водорослей монадная структура присуща клеткам, служащим для размножения – бесполого (зооспоры) и полового (гаметы). Кроме жгутиков, у монадных форм обычно развиваются органы фоторецепции (например, стигма или глазок) и сократительные (пульсирующие) вакуоли, выполняющие функцию осморегуляции. Монадный тип структуры преобладает среди эвгленовых, динофитовых, криптофитовых и золотистых, отмечается у зеленых и желтозеленых водорослей. У бурых и диатомовых в вегетативном состоянии этот тип структуры отсутствует, а появляется на стадии размножения. У красных водорослей монадные стадии не выявлены.
Амебоидный (ризоподиальный) тип морфологической организации наблюдается в основном у одноклеточных водорослей, лишенных твердой клеточной оболочки и постоянной формы тела (рис. 15; 27).
Рис. 27. Амебоидный (ризоподиальный) тип морфологической организации таллома у водорослей:
1 – Chrysamoeba;
2 – Myxochrysis;
3 – Myxochloris;
4 – Dinamoebidium
57
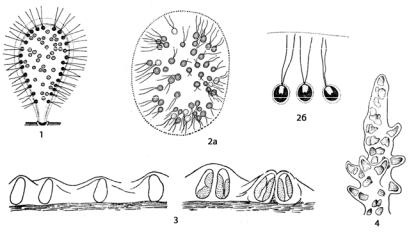
Такие организмы способны к амебоидному движению с помощью временно образующихся на поверхности клетки цитоплазматических выростов – псевдоподий (если выросты длинные и тонкие, то они называются ризоподиями). Иногда несколько клеток подобного строения соединяются своими ризоподиями и образуют колонии или сливаются в плазмодий – голую лишенную оболочки массу, содержащую несколько или много ядер. Многие амебоидные организмы ведут прикрепленный образ жизни. Этот тип структуры наблюдается у представителей немногих отделов, например, у золотистых (Chrysamoeba, Myxochrysis, рис. 27: 1, 2), желтозеленых (Myxochloris, рис. 27: 3), динофитовых
(Dinamoebidium, рис. 27: 4), а также у спермациев (мужских половых клеток) красных водорослей.
Рис. 28. Пальмеллоидный (гемимонадный) тип структуры вегетативного тела у водорослей: 1 – Apiocystis; 2 – Tetraspora
(а – общий вид колонии; б – часть колонии при большом увеличении); 3 – Chlorosaccus; 4 – Hydrurus
Пальмеллоидный (гемимонадный) тип структуры представлен неподвижными, одетыми оболочками или голыми клетками, имеющими черты монадной организации и погруженными в общую слизь (рис. 28). Такие слизистые талломы имеют разные размеры и очертания, встречаются у тетраспоровых водорослей из отдела зеленые (Apiocystis, Tetraspora), желтозеленых (Chlorosaccus) и золотистых (Hydrurus) водо-
рослей.
58
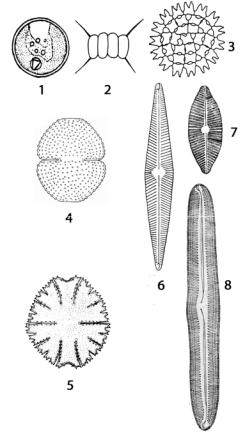
Коккоидный тип структуры объединяет одноклеточные, колониальные и ценобиальные водоросли, имеющие собственные оболочки, неподвижные в вегетативном состоянии (кроме некоторых диатомовых и десмидиевых) и полностью лишенные признаков монадной организации (рис. 29). Этот тип организации наиболее характерен для зеленых тре-
буксиевых – Chlorella, хлорококковых – Scenedesmus, Pediastrum, десмидиевых – Cosmarium, Micrasterias, диатомовых – Navicula, Pinnularia и
других.
Рис. 29. Коккоидный тип структуры таллома у водорослей:
1 – Chlorella;
2 – Scenedesmus;
3 – Pediastrum;
4 – Cosmarium;
5 – Micrasterias;
6, 7 – Navicula (разные виды); 8 – Pinnularia
59
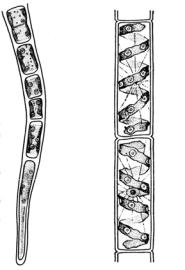
Нитчатый (трихальный) тип структуры представлен талломами, состоящими из клеток, соединенных в простые или разветвленные нити (например, зеленая водоросль Ulothrix). Клетки нити непрерывно делятся поперечными перегородками, за счет чего нить нарастает в длину (рис. 30). Нити могут быть свободноживущими, прикрепленными или
|
объединенными в слизистые ко- |
||||||
|
лонии. Клетки в нитях могут |
||||||
|
быть |
однотипными (зигнемовая |
|||||
|
водоросль Spirogyra) или, как, |
||||||
|
например, у Ulothrix, может фор- |
||||||
|
мироваться |
базальная |
клетка |
||||
|
клиновидной формы, бесцветная |
||||||
|
и служащая для прикрепления к |
||||||
|
субстрату, и вегетативные клетки |
||||||
|
цилиндрической формы с хло- |
||||||
|
ропластами. |
|
|
|
|
||
|
Разнонитчатая или гетеротри- |
||||||
|
хальная ступень |
морфологиче- |
|||||
|
ской дифференциации характери- |
||||||
|
зуется тем, что таллом состоит из |
||||||
|
системы нитей, стелющихся по |
||||||
|
субстрату, |
имеющих |
опорную |
||||
|
функцию и отходящих от них |
||||||
Рис. 30. Водоросли |
вертикальных нитей, выполняю- |
||||||
щих |
функцию |
ассимиляции и |
|||||
нитчатой структуры |
размножения. Нити могут быть |
||||||
(слева – Ulothrix, |
|||||||
однотипными |
или |
их |
строение |
||||
справа – Spirogyra) |
|||||||
может различаться (рис. 31). |
|||||||
|
|||||||
Примером водорослей с разнонитчатой структурой таллома может быть зеленая водоросль
Stigeoclonium (рис. 31: 1). У Draparnaldia (рис. 31: 2), многие виды кото-
рой являются эндемичными для озера Байкал, редуцированы горизонтальные нити, а вертикальные достигают большой сложности, разделяясь на центральную ось и боковые, с разной структурой клеток. У гетеротрихальной зеленой водоросли Coleochaete (рис. 31: 3) редуцирована вертикальная составляющая таллома, а горизонтальные веточки срастаются своими боковыми сторонами, образуя пластинки до 2 мм в диаметре. В онтогенезе водоросли иногда появляется вертикальная нить, состоящая из нескольких клеток, впоследствии отмирающая. Наиболее
60