

Основы альгологии и микологии Охапкин
.pdfРис. 92. Строение швов в панцире диатомовых водорослей: А – шов щелевидный (1 – разрез в области центрального узелка;
2 – шов в районе конечного узелка); Б – часть створки Surirella с каналовидным швом: нщ – наружная щель; вщ – внутренняя щель; к – канал, соединяющий наружную и внутреннюю щели; с – канал, соединяющий две ветви шва
в центральном узелке; кщ – конечная щель; в – конец внутренней щели шва
Размножение – вегетативное деление клеток надвое, имеется половой процесс. Перед делением клетки протопласт увеличивается в размерах, в нем накапливаются капли масла, при этом гипотека и эпитека раздвигаются, соприкасаясь лишь краями поясковых ободков. Ядро делится митотически, у многих диатомовых делению ядра предшествует деление хлоропластов. Каждый дочерний протопласт получает половину панциря материнской клетки, вторая половина (всегда гипотека) у обеих клеток достраивается заново (рис. 93).
171
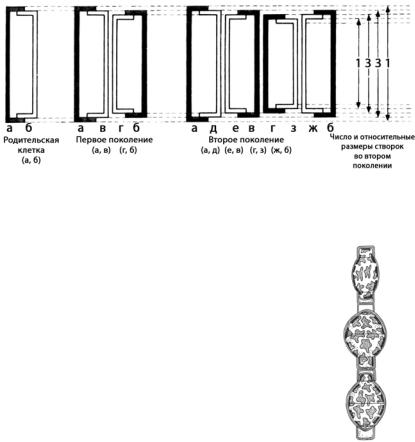
Рис. 93. Схема постепенного уменьшения размеров панциря диатомовых водорослей (вид с пояска)
врезультате серии последовательных клеточных делений
Всвязи с этим одна дочерняя клетка по размерам совпадает с материнской; размер второй дочерней клетки будет меньше примерно на толщину створки.
Участи видов диатомовых в результате многократ-
ных делений размер клеток может снижаться в три раза в сравнении с первоначальным.
Компоненты новой створки у каждой дочерней |
|
|
клетки формируются внутри особого окруженного |
|
|
мембраной пузыря, возникающего под плазмалем- |
|
|
мой, благодаря слиянию отдельных пузырьков Голь- |
|
|
джи. Мембрана, ограничивающая этот пузырь, назы- |
|
|
вается силикалеммой. В период формирования но- |
|
|
вых створок в клетках идет активный транспорт иона |
|
|
SiO32-, его полимеризация внутри пузыря до кремне- |
|
|
зема и формирование новой створки. Створка и поя- |
Рис. 94. |
|
сковый ободок новой дочерней клетки формируются |
||
Ауксоспоры |
||
в отдельных пузырях. При полном формировании |
Melosira |
|
недостающих частей панциря и их освобождении из |
|
пузыря часть силикалеммы, обращенная внутрь протопласта, становится частью плазмалеммы дочерней клетки, у некоторых плазмалемма дочерних клеток формируется заново без участия силикалеммы. Часть силикалеммы, обращенная кнаружи протопласта, возможно, составляет часть органического покрова, обрамляющего панцирь снаружи.
172
Уменьшению размеров клеток диатомовых водорослей в ходе вегетативного деления противостоит их увеличение в результате полового процесса, сопровождающегося образованием особых спор роста – ауксоспор (от греческого «ауксо» – увеличиваю, расту) (рис. 94).
Процесс измельчания клеток диатомовых водорослей предотвращается также в период прорастания так называемых покоящихся спор и
|
покоящихся |
клеток. |
У |
||
|
пеннатных |
диатомей |
|||
|
половой процесс |
напо- |
|||
|
минает |
конъюгацию |
|||
|
десмидиевых |
водорос- |
|||
|
лей, отличие заключа- |
||||
|
ется в том, что при об- |
||||
|
разовании гамет проис- |
||||
|
ходит |
редукция |
числа |
||
|
хромосом (гаметическая |
||||
|
редукция). |
|
|
|
|
|
При этом две клетки |
||||
|
сближаются и выделяют |
||||
|
общую слизь (рис. 95: |
||||
|
1–3). В каждой клетке |
||||
|
диплоидное ядро делит- |
||||
|
ся мейотически на че- |
||||
|
тыре |
гаплоидных, |
из |
||
|
которых у одних видов |
||||
|
отмирают три, а у дру- |
||||
|
гих – два ядра. |
|
|
||
|
В |
первом |
случае |
||
|
весь протопласт |
мате- |
|||
|
ринской клетки |
стано- |
|||
|
вится гаметой, во вто- |
||||
|
ром – формируется две |
||||
|
гаметы в каждой мате- |
||||
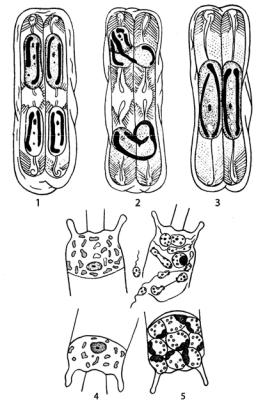
Рис. 95. Типы полового процесса |
ринской клетке. Если в |
||||
у диатомовых водорослей: |
каждой клетке образо- |
||||
1–3 – физиологическая |
вались по две гаметы, |
||||
анизогамия у Pinnularia; |
то одна из них перепол- |
||||
4, 5 – оогамия у Biddulphia |
зает в другую клетку и |
||||
|
сливается с |
неподвиж- |
|||
|
|
|
|
|
173 |
ной гаметой. Оставшаяся в первой клетке неподвижная гамета сливается с подвижной, переползающей из другой материнской клетки. Если в материнских клетках сформировалось по одной гамете, то одна из них – подвижная – переползает в другую клетку и сливается с неподвижной гаметой. В результате в первом случае формируются две зиготы и две ауксоспоры, во втором – одна. Половой процесс такого типа рассматривается как физиологическая анизогамия. У некоторых видов в каждой материнской клетке образуется по две гаметы, различающиеся по размерам. Затем гаметы разных размеров из разных материнских клеток попарно сливаются. У части видов известна автогамия (апогамия), когда ауксоспора образуется из одной материнской клетки. При этом диплоидное ядро исходной клетки претерпевает мейоз, два гаплоидных ядра отмирают, два оставшихся сливаются с образованием нового диплоидного ядра.
Половой процесс происходит быстро, одна или две образовавшихся зиготы интенсивно растут и превращаются в ауксоспоры определенной формы и строения, отличающиеся от вегетативных клеток. Ауксоспора в свою очередь прорастает в вегетативную клетку, имеющую большие размеры и свойственную виду структуру створок.
У центрических диатомовых водорослей обнаружен оогамный поло-
вой процесс (Melosira, Biddulphia) (рис. 95: 4, 5). Женская репродуктив-
ная клетка (оогоний) образует либо одну (Melosira varians), либо две (Biddulphia mobiliensis) яйцеклетки. Мужская репродуктивная клетка (сперматогоний) образует два – четыре (Melosira) или четыре (Biddulphia) сперматозоида, оплодотворяющих яйцеклетку. Сперматозоиды центрических диатомей – мелкие, подвижные клетки с одним опушенным жгутиком, без глазка. Процесс образования самих сперматогониев неодинаков у разных видов центрических диатомовых водорослей. У некоторых видов вегетативные клетки непосредственно становятся сперматогониями. У Melosira varians из одной вегетативной клетки в результате вегетативного деления возникает четыре – восемь более мелких с упрощенной структурой панциря и небольшим количеством хлоропластов сперматогониев. У Biddulphia mobiliensis после ряда митозов из вегетативной клетки формируются шестнадцать антеридиев, которые освобождаются после расхождения ее створок. Мейоз и образование сперматозоидов происходит свободно в воде. Яйцеклетки (у Melosira и Biddulphia) оплодотворяются внутри оогония, т.е. осуществляется истинная оогамия, у других центрических диатомей – только после выхода яйцеклетки из оогония, т.е. наблюдается примитивная оогамия. У
174
некоторых видов центрических диатомовых водорослей (например, Cyclotella meneghiniana) описана автогамия. Большой вклад в изучение и описание полового процесса диатомовых водорослей внесли русский ученый Б. М. Персидский и немец Г. Штош.
При неблагоприятных условиях диатомовые водоросли образуют покоящиеся клетки и покоящиеся споры. Покоящиеся споры отличаются по форме и структуре панциря от вегетативных клеток. Спора прорастает в вегетативную клетку, размеры которой превышают размеры самой споры.
В зависимости от конфигурации клетки и структуры створок у диатомовых обычно выделяют два класса.
Класс Centrophyceae – Центрические. Створки имеют радиальную симметрию, через них можно провести не менее трех плоскостей симметрии. Клетки одиночные или соединены в нитевидные или цепочковидные колонии. Форма панциря различная. Створки в очертаниях круглые, эллиптические, треугольные и многоугольные; их поверхность плоская или выпуклая, иногда вогнутая, реже волнистая. Шва нет. По краю створки расположены выступы в виде выростов, шипов, щетинок, при помощи которых клетки соединяются в колонии. Хлоропласты в виде многочисленных мелких зерен или одной или нескольких пластинок. Различные виды центрических диатомей широко представлены в планктоне Мирового океана, а также пресных континентальных водоемов – рек, озер, водохранилищ, где они являются одними из главных продуцентов органического вещества. Это очень древняя группа, представители известны еще с раннего мела; к настоящему времени многие роды целиком вымерли, из других родов сохранились только некоторые виды. Класс Centrophyceae делится на ряд порядков, из них наиболее крупные – Thalassiosirales, Melosirales, Coscinodiscales.
В планктоне морей широко распространены виды рода косцинодискус (Coscinodiscus) (рис. 96: 1). Клетки внешними очертаниями имеют сходство с цилиндром, диаметр которого часто меньше высоты. На створках радиальными или тангентальными (расположенными по хорде окружности панциря) рядами, реже в беспорядке располагаются ареолы, по краю створки имеются шипики.
175
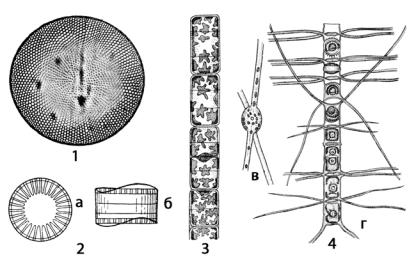
Рис. 96. Центрические диатомовые водоросли: 1 – Coscinodiscus (панцирь со створки); 2 – Cyclotella (а – вид со стороны створки, б – с поясковой стороны); 3 – Melosira; 4 – Chaetoceros (в – клетка со створки, г – часть колонии)
В морских и пресных водах, иногда на дне водоемов встречаются виды рода циклотелла (Cyclotella) (рис. 96: 2) либо в виде одиночных клеток, либо, реже, в виде колоний из рыхлых цепочек. Краевая зона створки несет радиальные штрихи или ребрышки, центр створки часто более выпуклый, у большинства видов бесструктурный. Хлоропласты мелкие, пластинчатые, расположены в периферическом слое цитоплазмы.
Цилиндрическую или боченковидную форму имеют панцири видов рода мелозира (Melosira) (рис. 94; 96: 3), обитающих в планктоне и бентосе пресных и соленых водоемов. Клетки соединены в плотные нити (колонии) своими створками при помощи слизи, иногда в скреплении створок принимают участие и шипики. Створки круглые, покрыты нежными гранулами. Створки соединяются своими расширенными загибами в середине клетки. Поясковые ободки появляются в период деления клетки. Загиб створки большой, покрыт беспорядочно расположенными мелкими гранулами. Хлоропласты многочисленные в виде пластинок, дисков или лопастные. Ниже представлен цикл развития центрической диатомовой водоросли на примере Melosira.
176
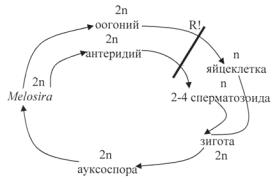
Цикл развития Melosira
У видов рода хетоцерос (Chaetoceros) (рис. 96: 4) формируются цепочковидные колонии, в которые объединяются клетки с помощью длинных полых щетинок или шипов, расположенных на краях створок. У ряда видов образуются вставочные ободки, увеличивающие объем клетки.
Класс Pennatophyceae – Перистые, или Пеннатные. Водоросли одноклеточные или колониальные. Со стороны пояска створки имеют двустороннюю симметрию, через них можно провести не более двух плоскостей симметрии. Панцирь симметричный по продольной оси, изопольный, реже гетеропольный, иногда дорсовентральный, редко асимметричный, с пояска линейный, таблитчатый, клиновидный, прямой, иногда S-образно изогнутый, часто с вставочными ободками и септами. Створки линейные, ланцетные, эллиптические, булавовидные, серповидные, овальные, яйцевидные и очень редко круглые, симметричные по продольной оси, изопольные или гетеропольные, реже симметричные по поперечной оси (рис. 91). По продольной оси створки проходит узкая бесструктурная полоса – осевое поле; в середине створки осевое поле часто расширяется, образуя центральное поле. У большинства видов вдоль середины осевого поля расположен щелевидный шов. Иногда шов сдвинут к краю створки или находится в киле (каналовидный шов). Хлоропласты в виде одной или нескольких крупных пластинок, реже – в виде многочисленных мелких пластинок.
Пеннатные диатомеи – пресноводные и морские водоросли, обитающие в бентосе на различных субстратах, и только единичные виды являются планктонными. Класспеннатныхдиатомеймоложе центрических.
177
Порядок Araphales – Бесшовные. Клетки одиночные или соединены в пучки, звездчатые или зигзаговидные колонии, в вегетативном состоянии неподвижны. Панцирь прямой. Створки от эллиптических до линейных, иногда булавовидные. Осевое поле от нитевидного до широколинейного, без шва.
У видов рода синедра (Synedra) (рис. 97: 1) клетки одиночные или соединены в пучковидные колонии. Панцирь с пояска палочковидный. Створки прямые, линейные, к концам суженные, с одним пластинчатым хлоропластом и тонкими, нежными параллельными штрихами, прерванными осевым полем. Со стороны пояска клетка имеет вид сильно вытянутого прямоугольника с двумя ребрами хлоропластов, тонкими, нежными штрихами, расположенными по загибу створки. Встречается в морских, солоноватых и пресных водоемах, в бентосе и обрастаниях, редко в планктоне.
Часто встречающиеся в пресноводных и морских водах представители рода астерионелла (Asterionella) (рис. 97: 2) – типичные компоненты планктона. Клетки соединены в звездчатые колонии (aster – звезда). Панцирь с пояска линейный, гетеропольный (базальный конец шире головного). Створки узколинейные, с головчатыми концами, головка на базальном конце крупнее, чем на свободном конце.
Виды рода фрагилярия (Fragilaria) (рис. 97: 3) напоминают Synedra, но соединяются створками при помощи шипиков в длинные лентовидные или загзагообразные колонии, обитающие в прибрежной зоне озер, прудов и других водоемов, среди высших водных растений, в обрастаниях, реже в планктоне. С пояска панцирь узкий, линейный. Форма створок от узколинейной до ланцетной, они часто расширены посередине, иногда волнистые или треугольные.
Род табеллярия (Tabellaria) (рис. 97: 4) объединяет широко распространенные пресноводные виды, часто встречающиеся в планктоне и бентосе различных водоемов с мягкой маломинерализованной водой с повышенной цветностью. Клетки соединены в звездчатые, лентовидные или зигзаговидные цепочки. Клетки с пояска имеют вид табличек, снабжены вставочными ободками и септами, створки линейные или овальные, на концах или на середине расширенные.
178

Рис. 97. Диатомовые бесшовные водоросли: 1 – Synedra (а – панцирь с пояска; б, в – со стороны створки); 2 – Asterionella (г – панцирь со створки;
д – общий вид колонии); 3 – Fragilaria (е – панцирь с пояска; ж, з – со створки); 4 – Tabellaria (и – общий вид колонии; к – панцирь со створки)
Порядок Raphales – Шовные. Это типичные обитатели бентоса пресных и морских водоемов, а также множество видов приурочено к сообществам обрастаний разных предметов, погруженных в воду, и высших водных растений. Клетки большей частью одиночные, подвижные, реже собраны в лентовидные или кустовидные колонии. Панцирь продольно или поперечно симметричный, створки от линейных до широко эллиптических, изредка S-образно изогнутые. Одна или обе створки имеют шов. Структура створки представлена штрихами, ребрами, которые располагаются поперечными рядами, иногда камерами.
Род пиннулярия (Pinnularia) (рис. 98: А–Д) – наиболее известный среди шовных диатомовых водорослей. Клетки одиночные, редко соединенные в ленты. Панцирь с пояска прямоугольный. Створки изопольные, линейные, линейно-эллиптические и ланцетные, с закругленными концами. Структура створок состоит из гладких поперечных ребер, представляющих собой полые камеры в толще панциря. Шов ните-
179
видный, центральный и конечные узелки хорошо развиты. Хлоропласты в виде двух пластинок, расположенных вдоль поясковых сторон клетки. Населяет преимущественно пресные водоемы, реже моря, часть видов – ископаемые. Со стороны створки видны ребра двух хлоропластов, а со стороны пояска – один пластинчатый. Смещение хлоропластов со створковой стороны на поясковую связано с появлением шва.
Виды рода навикула (Navicula) (рис. 98: Е) имеют много общего с видами Pinnularia, но отличаются от последней отсутствием камер в створках. Структура створки состоит из нескольких поперечных ребрышек, состоящих из ареол разной формы (округлых, эллипсоидных). Концы клеток более заостренные, суженные, по форме створки напоминают лодочку (navicula по-латыни – лодочка).
Виды гиросигмы (Gyrosigma) (рис. 98: Ж) имеют створки, по форме напоминающие букву S и имеющие штриховку панциря в виде мелкой сеточки. В прибрежной литоральной зоне и в обрастаниях главным образом в пресных водоемах широко распространены виды рода цимбелла (Cymbella) (рис. 98: З). Их створки полулунной формы, с прямым или вогнутым брюшным и выпуклым спинным краем. Шов обычно более или менее смещен к брюшному краю створки. Хлоропласт один, располагается с поясковой стороны. Некоторые виды прикрепляются к субстрату слизистой ножкой, выделяемой протопластом на одном конце створки, или размещаются в трубках из слизи, где клетки могут свободно передвигаться.
Из водорослей с каналовидным швом очень разнообразны в бентосе, обрастаниях и реже в планктоне водоемов виды рода нитцшия (Nitzschia) (рис. 98: И). Створки имеют вид палочек с заостренными концами. На каждой створке имеется особая продольная складка – киль с каналовидным швом, идущим вдоль каждой створки клетки.
Диатомовые водоросли – важнейший компонент биоты наземных и водных экосистем с всесветным распространением, широко представлены в разных местообитаниях, часто встречаются в массовом количестве. Много видов диатомовых водорослей обитают в почвах различных типов и природных зон. Диатомовые водоросли по некоторым оценкам образуют около четверти мировой первичной продукции, создаваемой растениями. Многие виды, имеющие шов, являются донными формами, обитающими в морских и пресных водах, другие (эпифиты) обрастают предметы, погруженные в воду, и растения. Бесшовные диатомеи играют огромную роль в планктоне морей, океанов и пресных вод.
180