

Основы альгологии и микологии Охапкин
.pdfБесполое размножение осуществляется за счет образования зооспор и апланоспор. Половое воспроизведение (хологамия, изогамия, анизогамия) обнаружено у немногих видов. После слияния гамет образуется подвижная зигота – планозигота. Имеются виды (Noctiluca scintillans) диплоидные в вегетативном состоянии и с гаметическим мейозом. Большинство видов динофитовых – гаплонты с зиготическим мейозом, у одного вида описан дипло – гаплофазный жизненный цикл. В неблагоприятных условиях у многих панцирных видов образуются толстостенные долговечные (до 16 лет) цисты, по форме напоминающие вегетативные клетки.
Некоторые специализированные паразитические формы динофитовых водорослей в период вегетативной фазы существуют как плазмодиальные организмы и теряют основные морфологические признаки монадных динофлагеллят. Они, как правило, размножаются зооспорами (диноспорами), очень похожими на обычных монадных представителей с двумя гетероморфными жгутиками, но со слабо развитыми поперечной и продольной бороздами.
Динофитовые водоросли обитают преимущественно в чистых пресных и соленых водах. Они встречаются обычно в холодное время года, известны криофилы, обитающие в снегу и придающие ему органогенную окраску. Развиваясь в массовых количествах, некоторые виды вызывают «цветение» воды, которое в морских водах называется красными приливами. Часто они бывают вызваны видами, вырабатывающими различные токсины, ядовитые для морских обитателей и человека.
В отделе Dinophyta выделяют либо один класс Dinophyceae, либо число классов возрастает до четырех. Из класса Desmophyceae большое значение в экосистемах морей имеет пророцентрум (Prorocentrum) (рис. 65: Д), иногда размножающийся в огромных количествах в прибрежной зоне морей и вызывающий «цветение» воды. Клетки более или менее овальные, с боков несколько сжатые. Тека состоит из двух блюдцевидных пластинок, сомкнутых вдоль краев клетки, за исключением переднего конца, гладких или украшенных шипиками или сетчатым рисунком. Борозд нет. Размножается путем продольного деления клетки, причем дочерние протопласты получают одну половину материнского панциря, а другую выделяют заново.
131
Класс Dinophyceae – Динофициевые – в соответствии с особенностями структурной организации особей делится на несколько порядков, из которых наиболее многочисленными являются порядки Gymnodiniales (гимнодиниевые) и Peridiniales (перидиниевые). Коккоидному уровню организации соответствует порядок Dinococcales, нитчатому – Dinotrichales с немногими представителями в каждом из них.
Порядок Gymnodiniales – Гимнодиниевые – отличаются от ос-
тальных динофитовых тонкой, нежной, гладкой или исчерченной текой. Панциря нет. Нередко имеются пузулы и стигма. Хлоропластов иногда нет. Формы фототрофные и гетеротрофные. Размножаются делением клетки, имеется пальмеллевидное состояние, известны цисты.
Для морских и пресноводных видов рода гимнодиниум (Gymnodinium) (рис. 65: А) характерна эллипсоидная, яйцевидная или шаровидная форма тела. Тека тонкая, нежная, гладкая или продольно исчерченная. Поперечная борозда экваториальная или слабо спирально закрученная, продольная борозда ограничена гиповальвой или заходит на эпивальву. Хлоропласты мелкие, буровато-коричневые или оливковозеленые.
Ночесветка (Noctiluca miliaris) (рис. 65: Б) является беспанцирной динофлагелятой, размер тела которой достигает 1–2 мм. Клетки бесцветные, без бороздок, с одним очень коротким продольным жгутиком. Возле жгутика имеется длинное хвостообразное поперечно-полосатое щупальце, функции которого – улавливать необходимые для питания организмы: одноклеточные водоросли, мелких животных. Цитоплазма содержит большое количество вакуолей с клеточным соком, ее тяжи простираются от периферии клетки к центральной массе, содержащей ядро, в постенном слое имеется большое количество липидных капель. Ночесветка – это один из организмов, вызывающих свечение моря. При массовом развитии ночесветки заметны простым глазом в виде светящихся точек, а вода при этом сильно фосфоресцирует голубоватым или серебристо-белым светом. Размножение осуществляется делением клеток или зооспорами, похожими на гимнодиниум, и отпочковывающимися от материнской особи. У некоторых видов известен половой процесс в виде изогамии, при образовании гамет, по-видимому, происходит мейоз (рис. 69).
В некоторых системах Noctiluca рассматривается как представитель класса Noctiluciphyceae, включающий исключительно морские организмы с крайне специализированным типом строения, сложным циклом развития и гетеротрофным типом питания.
132
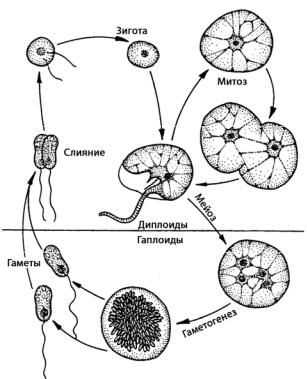
Рис. 69. Жизненный цикл Noctiluca scintillans
Порядок Peridiniales – Перидиниевые – характеризуется тем, что клетки этих водорослей имеют разнообразную форму, с поперечной и продольной бороздами и расположенными в них двумя жгутами. Оболочка в виде перипласта или панциря, состоит из щитков или пластинок. У большинства имеются хлоропласты. Продукты ассимиляции – крахмал или масло. Размножение делением в подвижном или неподвижном состоянии; плоскость деления обычно расположена косо. Новая клетка получает или половину материнской оболочки, или же дочерние клетки выходят из панциря и образуют новую оболочку. Описан половой процесс в форме хологамии и гаметогамии.
133
Многие представители обитают как в пресных, так и в морских водах, часто их развитие достигает степени «цветения» воды. Известны виды, вступающие в симбиоз с растениями и животными.
В морских и пресных водоемах часто встречаются различные представители родов перидиниум (Peridinium) и церациум (Ceratium) (рис. 65: В, Г). У перидиниума с брюшной и спинной сторон клетки овальных или округлых очертаний, сбоку они уплощенные. Клетки имеют мощный панцирь, состоящий из щитков, часто снабженных различными сосочками, шипиками, порами и другими скульптурными украшениями. Поперечная экваториальная бороздка выстлана щитками, которые образуют поясок, разделяющий клетку на почти равные части: верхнюю, апикальную эпивальву и нижнюю, антапикальную гиповальву. Швы у старых клеток достигают значительной ширины. Хлоропласты и глазок есть не у всех видов. При делении клеток дочерние особи образуют новый панцирь полностью, старый при этом разрушается. Известны зооспоры, похожие на гимнодиниума.
У видов рода церациум (Ceratium) клетки сильно вытянуты в продольном направлении и сильно сжаты дорсовентрально. Протопласт одет толстым панцирем. Эпивальва обычно продолжена в длинный прямой или изогнутый вправо апикальный рог с верхушечной порой на конце или без нее. Гиповальва с двумя-тремя рогами большей частью неодинаковой длины, из них рог, противолежащий апикальному, называется антапикальным, а остальные один-два рога – задними боковыми (правым и левым). Тело с вогнутой брюшной стороной и выпуклой спинной. Пластинки, слагающие панцирь, соединены между собой плотно, не образуя широких швов. Хлоропласты желто-бурые, многочисленные, постенные, дисковидные. Запасные питательные вещества – крахмал, жир, гликоген. Пластинки (щитки) покрыты системой нежных, расположенных в виде сетки складок, которые включают четырехшестиугольные или округлые углубления (ареолы).
Размножение в подвижном состоянии делением в косом направлении. В конце вегетационного периода образуются цисты. У одного вида описана анизогамия. Большинство видов морские, в пресных водах известны только три вида.
134
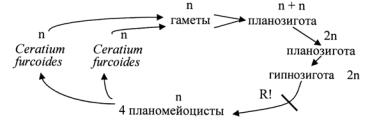
Цикл развития Ceratium
Развтие методов культивирования динофлагеллят позволило наблюдать у многих видов половой процесс. Переходу к половому размножению способствовало обеднение культуральной среды азотом. У многих видов наблюдали формирование голых изогамет, отличающихся от вегетативных клеток меньшими размерами и более светлой окраской. Часто отмечаются гомоталличные формы, т.е. половой процесс происходит из клеток одного клона. У других видов отмечен гетероталлизм и гетерогамия. Мужская гамета меньших размеров проникает в женскую, более крупную гамету, через отверстие поблизости от места прикрепления жгутиков. После слияния гамет образуется подвижная планозигота, передвигающаяся с помощью двух сохраняющихся задних жгутиков гамет, иногда остаются и передние жгутики. Период движения планозигот сопровождается их ростом, затем из них формируются одетые трехслойной оболочкой гипнозиготы (цисты). При их прорастании у всех изученных в этом отношении динофлагеллят происходит мейоз, и, таким образом, динофлагелляты – это гаплоидные организмы.
В современных системах эукариотных организмов отдел Dinophyta объединяют в одну большую группу альвеолата (Alveolata) вместе с типами инфузорий (Ciliophora) и споровиков (Apicomplexa). Это объединение основано на следующих ультраструктурных признаках: наличие трубчатых крист в митохондриях, наличие особо устроенного клеточного покрова, включающего у всех трех групп расположенные под плазмалеммой альвеолы, а также сходное строение стрекательных органелл. С современных позиций споровики и инфузории рассматриваются как вторично бесцветные, т.е. утратившие хлоропласты организмы.
135
йЪ‰ВО Haptophyta (Prymnesiophyta) – Й‡ФЪУЩЛЪУ‚˚В (ирЛПМВБЛУЩЛЪУ‚˚В) ‚У‰УрУТОЛ
å елкие, преимущественно морские свободноплавающие одноклеточные или колониальные, реже неподвижные коккоидные и пальмеллоидные организмы. Хлоропласты золотисто-бурого цвета, содержат хлорофилл а, с (с1 и с2) и каротиноиды с преобладанием фукоксантина. Имеется уникальный нитевидный орган – гаптонема, расположенный меж-
дудвумяизоконтными (иногда гетероконтными) жгутиками.
У большинства видов клетки имеют шаровидную или эллипсоидную форму, иногда клетки уплощены с боков. В жизненных циклах некоторых видов (род Pavlova) есть амебоидные стадии. Поверхность клетки покрыта органическими чешуйками или пластинками, расположенными в один или несколько рядов. У части видов слой более мелких чешуек покрывает снаружи гаптонему. У кокколитофорид на поверхности клеток размещаются крупные кокколиты. Форма их разнообразна, но часто встречаются округлые или дисковидные кокколиты. Кокколиты пропитываются карбонатом кальция в виде кальцита или арагонита. Некоторые кокколиты имеют внутренний канал. Чешуйки и некоторые виды кокколитов образуются в аппарате Гольджи, а затем транспортируются на поверхность клетки. Плотный покров коккоидных клеток и нитевидных форм состоит из нескольких слоев тесно упакованных чешуек и богат пектиновыми веществами. Гаптофитовые рода павлова не имеют типичных чешуек на поверхности клетки.
Монадные гаптофитовые имеют два жгутика равной или, реже, неравной длины, которые отходят от вершины переднего конца тела, у уплощенных форм – сбоку (рис. 70). Жгутики гладкие или у немногих покрыты тонкими волосками. Между ними расположена гаптонема – специфическая структура гаптофитовых, отсутствующая у других групп водорослей. От базальных тел жгутиков отходит сложно устроенная система корней, очень разнообразная у представителей разных порядков гаптофитовых. Гаптонема – это тонкая нить, у многих видов способная закручиваться спирально. Иногда гаптонема сильно редуцирована, имеет вид хоботка и заметна только в электронный микроскоп.
136

А |
Б |
Рис. 70. Особенности |
|
|
|
||
|
|
строения некоторых |
|
|
|
гаптофитовых: |
|
|
|
А – Prymnesium: |
|
|
|
1– жгутики; 2 – гаптонема; |
|
|
|
3 |
– хлоропласты; 4 – ядро; |
|
|
5 |
– аппарат Гольджи; |
|
|
6 |
– капли жира; |
|
|
7 |
– хризоламинарин; |
|
|
Б – Chrysochromulina |
|
Центр гапптонемы занят лентой из 6–8 микротрубочек, окруженных каналом эндоплазматической сети. Микротрубочки отходят от одной из кинетосом (базальных тел жгутиков), и поэтому многие авторы считают ее одним из корешков жгутика, хотя у самой гаптонемы обнаруживаются фибриллярные корешки. Функции гаптонемы различны: она может выполнять роль прикрепительной органеллы, участвовать в процессе движения клетки, а также участвовать в улавливании и поглощении клеткой твердых органических частиц (рис. 71).
Рис. 71. Схема захвата при помощи гаптонемы и заглатывания пищи на заднем конце клетки Chrysochromulina: 1–14 – последовательные этапы захвата и заглатывания пищевого объекта. Круговые стрелки указывают направление вращения клетки, прямые стрелки – движение частиц вдоль гаптонемы; б – бактерии; г – гаптонема; ж – жгутики
137
Вклетках содержится 1–2 хлоропласта с одним пиреноидом. Оболочка хлоропласта состоит из четырех мембран, каждая ламелла включает 3 тилакоида, опоясывающая ламелла отсутствует. Питание автотрофное, но есть и фаготрофные виды (хризохромулина – Chrysochromulina). Стигма имеется лишь у немногих видов (хорошо развита у видов рода павлова), находится внутри хлоропласта и состоит из одного ряда окрашенных в красный цвет глобул. Парафлагеллярное тело найдено только у одного вида. Сократительная вакуоль известна у пресноводных видов и расположена в передней части клетки. В периферическом слое цитоплазмы присутствуют многочисленные стрекательные органеллы, которые у гаптофитовых называются мукоцистами. Это шаровидные, веретеновидные или палочковидные тела, выделяющие наружу слизь в форме тонких нитей при раздражении клетки. В клетках можно наблюдать включения хризоламинарина и липидов. Ядро одно, с одним или несколькими ядрышками. Размножение осуществляется путем бинарного деления клеток в подвижном и пальмеллевидном состоянии. Половой процесс изучен недостаточно и известен в форме изогамии у немногих видов.
Гаптофитовые обитают в основном в морях и океанах; в солоноватых и пресных водах их немного. Кокколитофориды – это обитатели открытых зон морей и океанов, ряд видов развивается в зоне шельфа, часто вызывая «цветение» морских вод. «Цветение» воды, вызванное развитием токсичных видов из родов примнезиум (Prymnesium) (рис.
70:А) и хризохромулина (Chrysochromulina) (рис. 70: Б), приводит к гибели морских животных, особенно разных видов рыб. Известковые панцири ископаемых и современных кокколитофорид образуют мощные отложения известняков. Некоторые известковые породы (например, мел) на 50–75% состоят из кокколитов. Остатки вымерших кокколитофорид используют для определения геологического возраста известковых пород.
Вразных системах отдел Haptophyta содержит либо один класс Haptophyceae (Гаптофициевые) с признаками отдела и несколько порядков, либо число классов может быть больше. Примнезиевые (порядок Prymnesiales) – это одноклеточные, монадные, свободноплавающие водоросли. Клетки покрыты плазмалеммой, дорсовентральные с двумя изоконтными жгутиками и гаптонемой. Хлоропласты пристенные, боковые, без глазка, с пиреноидами или без них. Размножение – продольным делением клетки надвое в подвижном состоянии. Примнезиум ма-
138
ленький (Prymnesium parvum) (рис. 70: А) часто вызывает гибель рыбы в рыбоводных хозяйствах, вызывая токсичное «цветение» воды. Хризохромулина (Chrysochromulina, рис. 70: Б) имеет очень длинную гаптонему, примерно в пять раз длиннее жгутиков. В клетках содержатся двачетыре золотисто-желтых хлоропласта. Распространена в морях северных и южных широт, имеются и пресноводные виды, с развитием которых связаны образование запаха у воды и гибель водных животных.
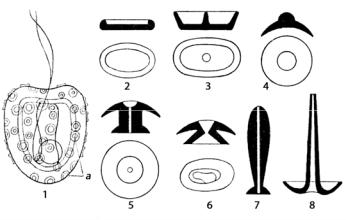
Кокколитофориды – порядок Coccolithophorales – это почти исключительно морская группа организмов, за исключением пресноводного гименомонаса розоватого (Hymenomonas roseola) (рис. 72).
Рис. 72. Hymenomonas roseola (1) и типы кокколитов кокколитофорид (2–8): а – кокколиты
Монадные клетки снабжены двумя гладкими одинаковой длины жгутиками, гаптонема хорошо развита не у всех видов. Кокколиты различной формы могут быть рассеяны по поверхности клетки со значительными промежутками, у других примыкают друг к другу, у третьих перекрываются (рис. 8; 72). Совокупность кокколитов в покрове клетки называется коккосферой. Форма и размеры кокколитов, строение коккосферы являются видоспецифичными и используются при идентификации кокколитофорид. В этом порядке известны виды, утратившие жгутики и ставшие неподвижными или же имеющие смену жгутиковой и прикрепленной стадий. Современные кокколитофориды населяют поверхностные слои (до глубины 150 м) Мирового океана. Большинство их обитает в морских бассейнах экваториальных и умеренных широт, в
139
холодных водах встречаются лишь немногие виды. Обилие кокколитофорид характерно не только для прибрежных вод, их много и в глубоководных, удаленных от берега районах. В ископаемом состоянии кокколиты известны с кембрия, но описаны и изучены только с юры.
Во многих системах водорослей гаптофитовые объединяют с золотистыми, так как они имеют существенное сходство с хризомонадами по структуре жгутиков, ультраструктуре хлоропластов, форме крист митохондрий, по типам хлорофиллов. Но наличие уникальной структуры – гаптонемы – позволяет ряду авторов выделять гаптофитовые в самостоятельный отдел. Хризомонадовые и гаптофитовые – филогенетически родственные группы, к которым близки диатомовые и бурые водоросли. Молекулярно-биологические данные свидетельствуют о достаточно большом расстоянии на филогенетическом древе между хризофитовыми и гаптофитовыми, что является дополнительным аргументом в пользу выделения гаптофитовых в самостоятельный отдел. В современной системе, предлагаемой протистологами, гаптофитовые формально отнесены к группе хромальвеолат (Chromalveolata) (биконты), поскольку они имеют своеобразную организацию клетки.
Следующие отделы водорослей, отличающиеся гетероконтными подвижными клетками, пластидами, иногда зелеными, но чаще за счет разнообразных каротиноидов имеющими окраску от желтых до коричневых, с хлорофиллами a и c и четырехмембранной оболочкой, в последние годы объединяются в группу Heterocontophyta или Ochrophyta, которой придается статус отдела.
йЪ‰ВО Raphidophyta – к‡ЩЛ‰УЩЛЪУ‚˚В ‚У‰УрУТОЛ
êафидофитовые водоросли – это небольшая четко обособленная группа одноклеточных фототрофных жгутиковых организмов, тело которых уплощено в дорсовентральном направлении. Их
хлоропласты содержат пигменты: хлорофиллы а и с (с1 и с2), β-каротин, антераксантин, лютеин и несколько ксантофиллов, у морских представителей отмечен фукоксантин. Основные вещества запаса – липиды.
Форма клеток рафидофитовых водорослей разнообразна: грушевидная, эллипсоидная, обратнояйцевидная, округлая до цилиндрической.
140