

Основы альгологии и микологии Охапкин
.pdfДомики бывают открытого, бокаловидного типа, как, например, у золотистой водоросли динобрион (Dinobryon), или закрытого, с небольшим отверстием для жгутика, как у эвгленовой водоросли трахеломонас (Trachelomonas). Домики могут быть сплошными или иметь различные перфорации, на их поверхности имеются разнообразные скульптурные украшения в виде шипов, бугорков, утолщений, точек. В одних случаях домики состоят только из органических веществ (Dynobryon), у других – пропитаны минеральными соединениями, например, солями железа (Trachelomonas). В отличие от клеточной оболочки, домик не участвует в процессе деления клетки.
У ряда водорослей целлюлозная оболочка не образуется, а покровы клетки устроены особым, часто специфическим образом. У эвгленовых водорослей под плазмалеммой расположен тонкий, плотный эластичный слой протопласта белковой природы толщиной до 70 нм, получивший название пелликулы. У большинства эвгленовых пелликула состоит из многочисленных полосок, начинающихся от глотки, раположенной на переднем конце тела, и по спирали огибающих клетку. Поверхность таких клеток продольно ребристая, ребра располагаются вдоль клетки. На поперечном разрезе ребра клетки имеют вид гребней с крючковатым изгибом, перемежающихся с глубокими бороздами (рис. 3).
Кроме того, пелликула может иметь разнообразные структуры в виде точек, бородавок, валиков и других. Если пелликула тонкая, тогда клетка может изменять свою форму при движении, толстая пелликула закрепляет форму клетки, и особи становятся неметаболичными.
Для криптофитовых водорослей характерен покров типа перипласта. Перипласт представляет собой плотное образование белковой природы, состоящее из двух слоев: один находится под плазмалеммой, а другой – над ней (рис. 4).
Внутренние и внешние пластинчатые структуры, из которых состоят эти слои, одинакового размера и толщины и тесно прилегают к плазмалемме. Как и пелликула у эвгленовых водорослей, перипласт криптофитовых состоит из серии полос, но направленных не по спирали, а прямыми продольными рядами от переднего конца клетки к заднему. Каждая полоса сформирована прямоугольными или шестиугольными пластинками, в углах которых расположены особые пузырьки, содержащие стрекательные органеллы, называемые эджектосомами. В местах контакта полос образуются борозды. Чередование гребней и борозд придает клетке рельефный контур. Перипласт обычно снабжен порами, через которые клетка сообщается с внешней средой.
31

У динофитовых клеточный покров получил наименование теки. Расположена тека под плазмалеммой и кроме белков содержит галактозу, арабинозу и галактуроновую кислоту. Основным элементом теки являются уплощенные пузырьки длиной от 600–800 нм до 0,8–1,2 мкм и толщиной 60–170 нм (рис. 5).
Рис. 5. Строение теки динофитовых водорослей Gymnodinium (1) и Glenodinium (2) по данным электронной микроскопии
Пузырьки располагаются под плазмалеммой и окружены мембраной. Число пузырьков варьирует от двух до нескольких сотен на клетку. Пузырьки либо заполнены зернистым веществом, либо содержат одиночные пластинки по одной в каждом пузырьке. Как правило, пузырьки располагаются в особом порядке, часто срастаются краями, образуя вокруг клетки прочный панцирь (рис. 6). В местах срастания пластинок формируются швы, являющиеся зонами роста панциря. На поверхности теки, вследствие своеобразия структуры пластинок возникают разнообразные узоры, иногда шипы. В теке, как и в перипласте, имеются поры.
Рис. 6. Строение теки (панциря) Peridinium по данным световой микроскопии: 1 – спинная сторона панциря; 2 – брюшная сторона панциря
У некоторых водорослей снаружи от плазмалеммы или клеточной оболочки формируются чешуйки, состоящие из органических (хитин, целлюлоза с гликаном) или минеральных (известковые, кремнеземные) веществ (рис. 7).
32
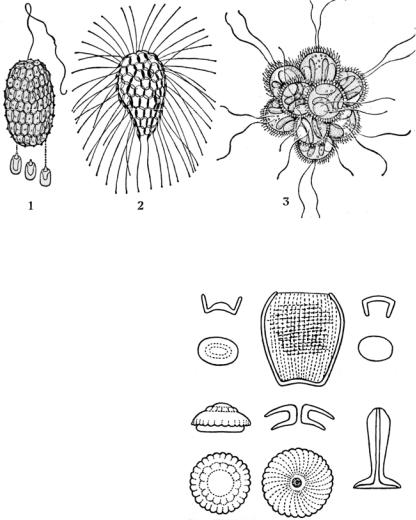
Рис. 7. Строение панциря золотистых водорослей: 1, 2 – Mallomonas (разные виды); 3 – Synura
Если у клетки нет оболочки, то чешуйки играют роль опорных элементов, поддерживают форму клетки и защищают ее от воздействий внешней среды. Часто чешуйки плотно упакованы в своеобразный панцирь. Известковые чешуйки
– кокколиты – наблюдаются преимущественно у морских водорослей кокколитофорид (рис. 8). Форма и расположение кремнеземных чешуек, встречающихся у золотистых водорослей, видоспецифичны, имеют диагности-
ческое значение.
Рис. 8. Форма кокколитов кокколитофорид
33

С клеточными покровами связаны своеобразные стрекательные (эджективные) структуры, выполняющие защитную функцию (рис. 9).
Рис. 9. Эджективные органеллы водорослей:
1 – Gonyostomum; 2 – Cryptomonas; 3 – эджектосома криптофитовой водоросли в распрямленном состоянии (в) и в процессе раскручивания (г);
4 – Gymnodinium; 5 – схема строения трихоцисты динофитовой водоросли в продольном (е) и поперечном (ж) разрезах; 6 – Peranema; а – эджективные органеллы; б – эджектосомы; д – трихоцисты; з – палочковидная органелла
Они напоминают по строению аналогичные структуры простейших и свидетельстуют о родстве водорослей и животных. Эти образования развиваются у подвижных представителей динофитовых, криптофитовых, эвгленовых, золотистых, рафидофитовых и зеленых водорослей. Как правило, они прикреплены к плазмалемме, пелликуле, перипласту или теке и имеют вид камер или пузырьков, окруженных одиночной мембраной. В пузырьках находятся свернутые спиралью стрекательные
34

нити, раскручивающиеся и выбрасывающиеся наружу при раздражении клетки или при наступлении неблагоприятных условий (например, токсическое воздействие). В деталях строение эджективных структур различается у водорослей разных отделов; у динофитовых они называются трихоцистами, у криптофитовых – эджектосомами, у золотистых – дискоболоцистами.
Клетки эукариотических водорослей содержат цитоплазму, основу которой составляет мелкогранулированный матрикс и ультраструктурные органеллы цитоплазмы: плазмалемму, эндоплазматическую сеть, аппарат Гольджи, микротрубочки, ядро и другие.
Цитоплазма клеток водорослей в большей или меньшей степени пронизана системой мембран, организованных в канальцы, цистерны и трубочки, и образующих единую эндоплазматическую сеть. Различают гладкую и шероховатую (за счет скопления рибосом) эндоплазматическую сеть, которая может встречаться в одной и той же клетке. Как и у других эукариот, эндоплазматическая сеть играет роль в передвижении и распределении веществ внутри клетки. Рибосомы, связанные с шероховатой эндоплазматической сетью, осуществляют синтез белка. Основной функцией эндоплазматической сети является полимеризация аминокислот до специфических полипептидов.
Аппарат Гольджи в клетке представлен совокупностью диктиосом, каждая из которых является системой собранных в стопки уплощенных мешочков, или цистерн, отграниченных элементарной мембраной (рис. 10). В одной диктиосоме может быть от трех-
семи до тридцати цистерн. От вздутых краев цистерн отделяются многочисленные мелкие пузырьки, или везикулы Гольджи. Число диктиосом в клетке водорослей может варьировать от одной до нескольких десятков. Функции диктиосом – это синтез из простых сахаров олигомерных углеводов, а также накопление, транспорт и секреция, главным
35
образом, полисахаридов, принимающих участие в образовании клеточных покровов. В цистернах аппарата Гольджи откладывается кремнезем и образуется новая створка у диатомовых водорослей, формируются элементы клеточного покрова золотистых, примнезиевых и некоторых зеленых водорослей – чешуйки. Путем эктоцитоза готовые чешуйки из пузырьков Гольджи перемещаются на периферию клетки и формируют клеточный покров. Так же формируются кокколиты у кокколитофорид. У рафидофитовых аппарат Гольджи принимает участие в формировании сократительных вакуолей. Аппарат Гольжди связан с секреторной деятельностью клетки, при его участии, по-видимому, образуются трихоцисты, эджектосомы, тека и другие элементы покрова клетки.
Эндоплазматическая сеть и аппарат Гольджи принимают участие в формировании вакуольной системы клетки. С возрастом число и размеры вакуолей увеличиваются. Вакуоли отделены от цитоплазмы мембраной – тонопластом, имеющим ту же природу, что и плазмалемма. В старых клетках вакуоли нередко сливаются в одну крупную центральную вакуоль. В подвижных клетках, а иногда и неподвижных, формируются сократительные пульсирующие вакуоли, выполняющие осморегуляторную функцию. У динофитовых и рафидофитовых водорослей аналогичную роль выполняют пузулы – особые вакуольные структуры, не способные к сокращению.
Помимо каналов эндоплазматической сети клетки эукариотических водорослей содержат микротрубочки, образованные белками тубулинами, по аминокислотному составу напоминающими актин мышц. Цитоплазматичесие трубочки различаются по положению в клетке и назначению: одни участвуют в образовании ядерного веретена, другие связаны с цитокинезом – участвуют в образовании фрагмопласта и фикопласта, третьи собственно цитоплазматические микротрубочки представляют собой цитоскелетную систему клетки. Особенно велика роль микротрубочек как элемента цитоскелета голых клеток. Кроме микротрубочек в цитоплазме клеток обнаружено большинство известных в настоящее время классов цитоплазматических частиц, окруженных одиночной мембраной и заполненных содержимым, отличающимся по электронной плотности: лизосомы, пероксисомы, сферосомы, физоды, ирисовые тела, ломасомы. Они различаются по своему химическому составу и функциональной роли в клетке.
Фотосинтетический аппарат является специфическим образованием растительных клеток, обеспечившим переход организмов на фотоавтотрофный способ питания. Состав фотосинтетических пигментов – один
36
из важнейших признаков, на основе которого строится деление водорослей на отделы. Так Cyanoprokaryota отличаются от остальных оксигенных фотосинтетиков тем, что фотосинтетический аппарат включает в основном только хлорофилл а, в то время как у Chlorophyta и Euglenophyta выявлены хлорофиллы а и b, у Chrysophyta, Dinophyta, Xanthophyta, Bacillariophyta, Cryptophyta, Raphidophyta и Phaeophyta – хлорофиллы
а и с, у Rhodophyta – хлорофилл а. Только у цианопрокариот, красных и криптофитовых водорослей обнаружены фикобилиновые пигменты – фикоцианин и фикоэритрин. Фикобилипротеины входят в состав особых гранул – фикобилисом, упорядоченно расположенных на поверхности фотосинтетических мембран. Значительные различия на уровне отделов выявлены также и в составе каротиноидов – каротинов и ксантофиллов.
Фотосинтетический аппарат клетки эукариотических водорослей представлен структурно оформленной органеллой – хлоропластом. Хлоропласты водорослей в отличие от хлоропластов высших растений очень разнообразны по числу, форме и размерам. Их количество колеблется у разных представителей от одного (Chlamydomonas, Dunaliella) до нескольких сотен (некоторые виды Euglena). Чем больше хлоропластов в клетке, тем обычно они меньше. По форме хлоропласты бывают чашевидными, пластинчатыми, корытовидными, сетчатыми, поясковидными, звездчатыми, линзовидными, спиралевидными и другой формы (рис. 11). В клетке они могут располагаться либо центрально, и тогда называются осевыми, или ближе к клеточной стенке – париетальные. Эти признаки очень изменчивы и сильно зависят от факторов внешней среды, изменяясь в процессе онтогенеза. У ряда зеленых сифоновых водорослей помимо хлоропластов обнаружены амилопласты – пластиды из группы бесцветных лейкопластов.
Хлоропласты глаукофитовых, красных и зеленых водорослей окружены двухмембранной оболочкой. У других водорослей хлоропласт снаружи окружен дополнительными одной или двумя мембранами хлоропластной эндоплазматической сети. У эвгленовых и большинства динофитовых присутствует одна мембрана хлоропластной эндоплазматической сети, и оболочка хлоропласта состоит из трех мембран. У криптофитовых, рафидофитовых, гаптофитовых, золотистых, желтозеленых, диатомовых, бурых, хлорарахниофитовых водорослей оболочка хлоропласта имеет четыре мембраны, так как у них присутствует дополнительтно две мембраны хлоропластной эндоплазматической сети. У водорослей с четырехмембранной оболочкой наружная мембрана хлоро-
37
пластной эндоплазматичсекой сети чаще всего переходит в наружную мембрану ядра.
Рис. 11. Основные формы хлоропластов у водорослей:
1 – чашевидный у Chlamydomonas; 2 – чашевидный двулопастный; 3 – чашевидный многолопастный у разных видов рода Arachnochloris; 4 – корытовидный у Monodus; 5 – дисковидные у Tribonema utriculosum; 6 – кольцевидные у Ulothrix; 7 – лентовидные у Tribonema vermichloris;
8 – спиралевидные у Spirogyra; 9 – сетчатый у Oedogonium; 10 – звездчатые осевые у Zygnema; 11, 12 – пластинчатый осевой у Mougeotia
38

Рис. 12. Схема расположения тилакоидов в хлоропластах водорослей: А – модель хлоропласта красной водоросли; Б – одиночное расположение
тилакоидов в хлоропластах красных водорослей; В – двухтилакоидные ламеллы криптофитовых; Г, Д – трехтилакоидные ламеллы в хлоропластах бурых
идинофитовых водорослей; Е – граны зеленых водорослей; ох – оболочка хлоропласта; т – тилакоиды; от – опоясывающий тилакоид; г – граны
Под оболочкой хлоропласта располагается тонкозернистая белковая основа – строма или матрикс, в который заключены уплощенные, одетые мембранами мешочки или пузырьки – тилакоиды или диски, содержащие фотосинтетические пигменты. В белковой строме расположены пластидные ДНК, РНК, метаболические гранулы, микротрубочки и рибосомы. У красных водорослей тилакоиды лежат в матриксе поодиночно, у криптофитовых – соединены по два, у золотистых, желтозеленых, диатомовых, бурых, динофитовых и эвгленовых – по три (рис. 12). У эвгленовых водорослей иногда может быть более трех тилакоидов в ламелле. У зеленых водорослей число тилакоидов в ламеллах колеблется от двух до шести, иногда до двадцати, в таких случаях стопки тилакоидов тесно прижаты друг к другу, и эти стопки называются гранами. В хлоропластах золотистых, желтозеленых, диатомовых и бурых водорослей с трехтилакоидными ламеллами находятся еще так называемые периферические (опоясывающие) ламеллы. Они располагаются параллельно оболочке хлоропласта и окружают остальные ламеллы, пересекающие хлоропласт. Хлоропласты разных водорослей содержат элек-
39

тронно-прозрачные участки, содержащие фибриллы ДНК – генофоры. Впервые генофоры были обнаружены в 1962 г. у хламидомонад.
В клетках прокариотических оксигенных фотосинтетиков морфологически оформленных хлоропластов нет. Фотосинтетичский аппарат у них представлен системой одиночных тилакоидов, которые пронизывают всю цитоплазму и располагаются параллельными рядами, опоясывающими периферическую часть клетки.
Рис. 13. Схема строения пиреноида красной водоросли: а – обкладка пиреноида; б – строма пиреноида; в – тилакоиды пиреноида; г – тилакоиды хлоропласта
Хлоропласты эукариотических водорослей содержат особые включения – пиреноиды (рис. 13). Это свойственные только водорослям и некоторым мхам тельца белковой природы, в основном состоящие из фермента рибулезобисфосфаткарбоксилазы (РУБИСКО). Пиреноиды принимают участие в синтезе различного рода запасных соединений, например, у зеленых водорослей – крахмала. Пиреноиды состоят из основного вещества – стромы, нередко погруженной в него системы ламелл и часто окружающей его обкладки. Строма пиреноида отличается от стромы хлоропласта более высокой плотностью и гомогенностью. На границе стромы хлоропласта и стромы пиреноида мембрана не обнаружена. Условно границей между пиреноидом и хлороплатом служит сильно преломляющая свет обкладка, которая может быть сплошной или состоять из отдельных фрагментов. Благодаря наличию обкладки пиреноид хорошо заметен даже в живых клетках. Обкладка пиреноида у зеленых водорослей состоит из крахмала, у красных – из особой формы крахмала – багрянкового крахмала, у эвгленовых – из парамилона, у бурых – из ламинарина, у золотистых откладываются липидные капли. Иногда у желтозеленых, некоторых эвгленовых, зеленых пиреноиды не имеют обкладки и называются голыми.
40