

Основы альгологии и микологии Охапкин
.pdfРазмножение вегетативное – фрагментацией таллома, реже акинетами. Собственно бесполое и половое размножение изучено недостаточно и у немногих представителей. Сифонокладовые в основном морские водоросли, распространенные в тропиках и субтропиках, реже – пресноводные, развивающиеся в водоемах разного типа, в бентосе и обрастаниях.
Широко распространены в морских и пресных водоемах виды рода кладофора (Cladophora) (рис. 35; 118).
Рис. 118. Внешний вид таллома (слева), выход зооспор из спорангия (в середине) и схема строения клетки Cladophora: 1 – пиреноид; 2 – ядро; 3 – хлоропласты
Таллом кладофоры нитчатый, обильно ветвящийся, состоящий из вытянутых цилиндрических или слегка вздутых к вершине клеток или сегментов. Клетки одеты толстой слоистой никогда не ослизняющейся оболочкой. Под оболочкой находится постенная цитоплазма с многочисленными ядрами и мелкими неправильной формы удлиненными, смыкающимися своими концами в постенную сеть хлоропластами. Совокупность фрагментов хлоропластной системы сегмента кладофоры образует как бы единую сетчатую пластинку с многочисленными пиреноидами. Молодые талломы прикреплены к субстрату ризоидами, в бо-
221
лее зрелом возрасте они отрываются, разрастаются в толще воды, образуя скопления «тины» грязно-зеленого цвета, не слизистые на ощупь.
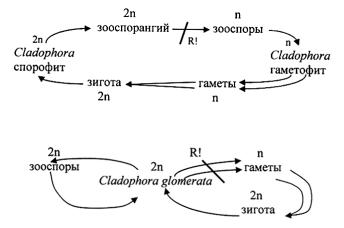
При бесполом размножении в конечных густо-зеленых сегментах ветвей образуются двужгутиковые или четырехжгутиковые зооспоры, которые выходят в воду из бокового отверстия в оболочке. Половой процесс – изогамия, гаметы двужгутиковые, мельче зооспор. Циклы развития в пределах рода отличаются. У широко распространенной в пресных водоемах европейской части России Cladophora glomerata талломы диплоидные, редукция гаметическая. Бесполое размножение осуществляется посредством диплоидных зооспор, прорастающих в диплоидные талломы. Многие морские виды имеют изоморфную смену форм развития со спорическим мейозом, а некоторые североамериканские виды гаплоидны.
Цикл развития морских представителей рода Cladophora
Цикл развития Cladophora glomerata
Ризоклониум (Rhizoclonium) (рис. 119) отличается от кладофоры очень слабым ветвлением, иногда таллом почти совсем не ветвится. У основания слоевища образуются короткие одноклеточные выросты – ризоиды. Клетки длинные, цилиндрические с немногочисленными (от двух до восьми) ядрами. Различные виды рода встречаюется как в пресных, так и морских водоемах вместе с кладофорой.
222
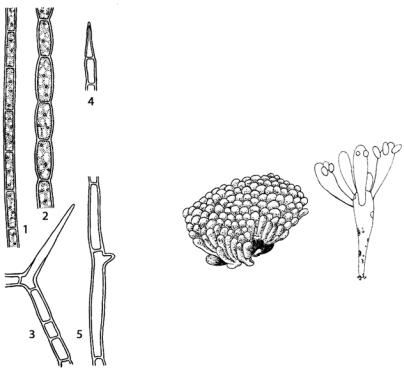
Рис. 119. Rhizoclonium hieroglyphicum: 1, 2 – части нитей с клетками различной формы; 3, 5 – ветвление талломов;
4 – терминальная клетка
а б
Рис. 120. Внешний вид таллома
Valonia (а) и пузырь
с маргинальными клетками (б)
Для некоторых морских видов установлен жизненный цикл с изоморфной сменой форм развития.
Очень своеобразное слоевище имеет морская водоросль валония
(Valonia) (рис. 120).
Это растение в разных системах относят либо к порядку Cladophorales, либо выделяют особый порядок Valoniales в связи с особым механизмом деления клеток.
У этого рода многоклеточность в онтогенезе возникает поздно в результате отчленения от старых клеток более молодых, которые значительно меньше по размерам. Большинство видов этого рода населяет тропические и субтропические моря. Таллом валонии представляет собой пузыревидное образование с более узкой базальной частью сифонального строения и прикреплен неклеточными ризоидами к грунту.
223
Позднее в любом месте таллома у его поверхности закладываются изогнутые перегородки в виде часового стекла, отчленяющие мелкие линзовидные сегменты. Эти клетки-сегменты называются маргинальными (краевыми). У основания пузыря маргинальные сегменты имеют очень небольшие размеры и нередко вырастают во вторичные ризоиды, часто с лопастными окончаниями. У просто устроенных видов валонии развитие таллома на этом заканчивается. У других видов из маргинальных сегментов, расположенных ближе к вершине слоевища, вырастают новые пузыри, которые в свою очередь могут формировать пузыри третьего порядка. Таким образом, вегетативное тело таких валоний представляет собой систему пузыревидных сегментов, развивающихся друг на друге.
Бесполое размножение у валонии происходит зооспорами, половой процесс – гетерогамия; редукция числа хромосом идет перед образованием гамет. Валонии с крупным, до нескольких сантиметров в длину, талломом предсталяют собой удобный объект для изучения биоэлектрических явлений, процессов проницаемости и аккумуляции веществ в клетках. Водоросль обладает хорошей способностью к регенерации: таллом полностью восстанавливается, еслисохранилосьхотябыодноядро.
Представители класса Bryopsidophyceae – Бриопсидофициевые
(Сифоновые) – это водоросли сифональной структуры, таллом которых не имеет радиальной симметрии. Слоевища разнообразного внешнего облика, часто простые или перисто разветвленные, иногда сложно переплетенные, образующие определенную морфологическую форму (подушковидную, цилиндрическую или другую). В большинстве случаев таллом хотя бы частично имеет вид нити. Часто слоевища могут достигать крупных размеров и значительной внешней дифференцировки, формально представляя собой одну гигантскую клетку, одетую толстой оболочкой. Тонкий постенный слой цитоплазмы находится между клеточной стенкой и крупной непрерывной вакуолью клеточного сока. В составе клеточной стенки наряду с целлюлозой встречаются маннан и ксилан. В цитоплазме располагаются многочисленные ядра и дисковидные хлоропласты, в состав пигментов которых, наряду с хлорофиллами и каротиноидами, входят два специфических ксантофилла – сифонеин и сифоноксантин. У ряда представителей наряду с хлоропластами известны бесцветные амилопласты. Таллом лишен перегородок, только в основании вегетативных ветвей и гаметангиев могут возникать перегородки с порой посередине, частично отделяющие их от таллома.
Размножение вегетативное фрагментацией и почками вегетативного размножения – пропагулами. В основном у этих водорослей встречается
224

половое размножение, бесполое (зооспорами) известно у немногих представителей. Половое размножение осуществляется при помощи двужгутиковых изогамет или гетерогамет, которые образуются в любой части таллома или в специальных гаметангиях, отделенных от таллома перегородками. Для большинства представителей жизненные циклы не ясны, у части представителей обнаружена гетероморфная смена форм развития. Предположение о наличии у большинства сифоновых диплоидного жизненного цикла с гаметическим мейозом в последнее время подвергается сомнению. Полагают, что мейоз может иметь место при прорастании зиготы в макроскопический таллом. Подавляющее большинство бриопсидофициевых встречается в субтропических и тропических морях, лишь немногие растут в Черном море и севернее; некоторые обитают в пресных водоемах умеренных широт или на влажной почве вблизи водоемов.
Виды порядка Bryopsidales – Бриопсидовые – не имеют амилопластов, служащих для образования крахмала. У дербезии (Derbesia) (рис. 121: 1–3), встречающейся в тропических морях и морях умеренных широт, таллом имеет вид стелющихся по субстрату и укореняющихся с помощью ризоидов сифоновых нитей.
Рис. 121. Строение бриопсидовых: 1–3 – Derbesia (1 – внешний вид слоевища, 2 – зооспора, 3 – зооспорангий); 4–6 – Halicystis (4 – первичные нити,
5 – молодые вертикальные нити, 6 – зрелый пузырь); 7–10 – Bryopsis (7 – веточка, 8 – ризоиды у основания веточек, 9 – часть слоевища с гаметангиями,
10 – перегородка в основании гаметангия в продольном сечении)
225
От них вверх отходит система вертикально растущих обычно ветвящихся нитей. Размножение у дербезии происходит путем образования 8–20 одноядерных стефаноконтных зооспор в овальных спорангиях. При формировании спорангиев концы вертикальных нитей вздуваются
иотделяются от остального таллома перегородками.
Уморской водоросли халицистис (Halicystis) (рис. 121: 4–6) таллом в виде шаровидного пузыря до 3 см в диаметре, в нижней части переходящего в горизонтальную часть, стелющуюся по субстрату и проникающую в субстрат. При развитии халицистиса сначала формируются горизонтальные нити на субстрате, затем они проникают внутрь известковых водорослей (например, литотамниона – Lithothamnion), на которых поселяется халицистис. Затем от горизонтальных нитей вверх отрастают короткие вертикальные побеги, на концах которых и формируются пузыри. Таким образом, слоевище халицистиса является дальнейшим усложнением таллома дербезии, а пузыри – это видоизмененные вертикальные побеги. Ежегодно осенью пузыревидная часть таллома отмирает, а зимует горизонтальная составляющая слоевища. Весной от нее вверх отрастают новые пузыревидные расширения.
Вегетативное размножение халицистиса осуществляется путем образования на материнском пузыре дочерних, последующим их отделением от таллома и «укоренением» на субстрате. Кроме того, на горизонтальной части таллома образуются новые пузыри, впоследствии отделяющиеся от материнского и переходящие к самостоятельному существованию. Кроме вегетативного размножения, у халицистиса известна гетерогамия. На мужских пузырях образуются желтовато-оранжевые плодущие участки, в которых формируются микрогаметы, на женских – зелёные участки, дающие начало макрогаметам. Созревшие гаметы с силой выбрасываются из пузыря через одну или несколько пор, возникающих в стенке над плодущими участками.
Вусловиях культуры было показано, что Halicystis parvula и Derbesia tenuissima являются соответственно гаметофитом и спорофитом одного
итого же организма, обнаруживающего гетероморфную смену форм развития. В результате слияния анизогамет халицистиса образуется диплоидная зигота, прорастающая нитями дербезии. Стефаноконтные зоо-
споры Derbesia tenuissima вырастают в пузыри Halicystis parvula.
Убриопсиса (Bryopsis plumosa) (рис. 34: 4; 121: 7–10), широко рас-
пространенного в Черном море на каменистых субстратах и небольших глубинах, таллом состоит из малоразветвленного ползучего «корневища», прикрепленного к субстрату ризоидами. От него вверх отходят
226
более или менее толстые нити, которые в верхней части перисто рассечены на боковые веточки, т.е. ветви расположены супротивно в одной плоскости; причем верхние короче нижних. Все это делает растение похожим на перо птицы. В основании боковых веточек образуются перетяжки, и клеточная стенка заметно утолщается. Перегородок в талломе нет до момента образования гаметангиев. Из основания вертикальных осей могут вырастать горизонтальные нити, стелющиеся по субстрату. От этих горизонтальных осей могут образовываться новые вертикальные.
Вегетативное размножение у бриопсиса осуществляется путем отделения от основной части таллома вертикальных веточек, которые у основания закупориваются и образуют ризоиды (рис. 121: 8), а затем укореняются и вырастают в самостоятельные растения. При поранении таллома часть протопласта отделяется от материнского растения, округляется, синтезирует новую оболочку и затем развивается в новую особь. Половой процесс – гетерогамия. Мужские гаметы с редуцированным хлоропластом, светло-желтые, без глазка и мельче женских. Женские гаметы с хлоропластом и глазком имеют светло-зеленую окраску. У многих видов бриопсиса мужские и женские гаметы развиваются в разных гаметангиях, в которые превращаются отдельные боковые веточки вертикальной составляющей таллома (рис. 121: 9). Боковые оси отделяются от главной оси перегородками (рис. 121: 10), но это бывает не всегда, поскольку гаметы могут сформироваться и в центральной главной оси. У других видов мужские гаметы образуются у конца гаметангия, а женские – у его основания. После копуляции гамет зигота сразу прорастает в мало ветвящуюся сифоновую нить – протонему, содержащую одно крупное ядро. В зависимости от условий произрастания в дальнейшем протонема может формировать новое растение бриопсиса (гаметофит) или же ее содержимое распадается на многочисленные стефаноконтные зооспоры, которые после выхода из протонемы прорастают в растение бриопсиса. Таким образом, формируется двухфазный цикл с гетероморфной сменой форм развития (бриопсис – гаметофит, протонема – редуцированный спорофит). Оба типа развития протонемы (сразу в гаметофит или через стадию зооспор) был выявлен у Bryopsis plumosa.
У Bryopsis halimeniae зигота прорастает нитчатой сифоновой стадией, описанной как отдельный вид – Derbesia neglecta. При прорастании зооспор у Derbesia neglecta (спорофит) образуются растения Bryopsis halimeniae (гаметофит).
227
Место мейоза в жизненном цикле бриопсиса остается спорным. Ранее считали, что редукция числа хромосом происходит при образовании зооспор (спорический мейоз), но в 1980-х гг. при изучении Bryopsis plumosa и B. hypnoides выяснилось, что мейоз имеет место при образовании гамет и гаметофит находится в диплоидной фазе так же, как и спорофит. Таким образом, в данном случае морфологические фазы жизненного цикла не связаны с ядерными.
В целом, у сифоновых водорослей в циклах развития обнаруживается у одних видов постепенная редукция спорофита, а у других – гаметофита. Исходным типом, по-видимому, можно считать близкую к изо-
морфной смену форм развития Bryopsis halimeniae и Derbesia neglecta. У Bryopsis plumosa и B. hypnoides постепенно редуцируется спорофит: при двухфазном цикле у них имеется спорогенная стадия проростка, при однофазном спор не образуется, а редуцированный спорофит (протонема) прорастает гаметофитом. У Derbesia marina гаметофит сильно реду-
цирован до пузыревидной стадии – Halicystis ovalis, а у D. lamourouxii
гаметофита нет вообще. Эти процессы обособления одной из фаз первоначально двухфазного жизненного цикла путем редукции или полной утраты другой фазы могли привести к постепенному становлению независимых родов Derbesia и Bryopsis. Таким образом, жизненные циклы бриопсидовых характеризуются большой сложностью, сопровождающейся нерегулярной, строго не фиксированной сменой форм развития.
У кодиума (Codium) (рис. 34: 3; 122) слоевище имитирует псевдопаренхиматозное строение.
Рис. 122. Внешний вид таллома Codium (слева), часть продольного среза через слоевище (в середине) и пузырь с гаметангиями (справа)
228
Макроскопические талломы (шаровидные, цилиндрически разветвленные, длиной до нескольких метров) возникают путем плотного переплетения простых трубок типично сифонового строения. В слоевище имеются две зоны: внутренняя, образованная пучком тонких бесцветных ветвящихся нитей, и наружная, состоящая из периферических нитей, от которых отрастают широкие и короткие пузыри (кортикальные пузыри или утрикулы), формирующие коровый слой таллома. Внутренний пучок нитей выполняет функцию проводящей системы. Строение и размеры кортикальных пузырей видоспецифичны. От верхней части утрикул нередко отходят бесцветные длинные легко отламывающиеся волоски. Они увеличивают общую поверхность таллома и способствуют росту интенсивности обменных процессов. В пузырях, играющих роль ассимиляторов, имеется много хлоропластов.
Размножение у кодиума вегетативное и половое. Двужгутиковые гаметы образуются в специальных гаметангиях – выростах кортикальных пузырей, отделяющихся от них перегородкой. Как и у многих других представителей порядка, у кодиума наблюдается гетерогамия. Мужские гаметангии окрашены в золотисто-желтый цвет, женские – темнозеленые. Зигота без периода покоя прорастает в новое растение. Кодиум является диплонтом; мейоз происходит при образовании гамет. В Черном море на глубине десять метров и более встречается Codium tomentosum. Этот вид может быть как двудомным, так и однодомным, причем в последнем случае мужские и женские гаметы возникают не только на одном растении, но и в одном и том же пузыре.
Клетки представителей порядка Halimedales – Халимедовые – имеют как хлоропласты, так и амилопласты. Клеточные стенки не содержат маннана. У каулерпы (Caulerpa) (рис. 123) слоевища сильно рассеченные, состоящие из распростертого шнуровидного или цилиндрического «корневища» до одного метра длиной, прикрепляющегося к грунту многочисленными ризоидами. От горизонтальной части вверх растут вертикально восходящие ассимилирующие побеги. В исходном случае это простые неправильно ветвящиеся нити. У других видов отходящая от цилиндрического корневища вертикальная ось образует уплощенные ассимиляторы, расположенные либо мутовками, как у хвощей, либо перисто в два ряда, напоминая побеги тиса. Ассимиляторы могут быть плоскими, листовидными на коротких стебельках, отрастающими сразу от горизонтальной части таллома. На поверхности первичных ассимиляторов впоследствии развиваются новые вторичные. Как и у других сифоновых, у каулерпы широко распространено вегета-
229

тивное размножение. При отмирании старых участков горизонтальной составляющей таллома его отдельные части с вертикальными ассимиляторами становятся отдельными дочерними растениями (вегетативное размножение).
Рис. 123. Внешний вид (1, 2) и продольный разрез таллома (3) Caulerpa
Перегородок в таком крупном макроскопическом талломе нет. Постенная цитоплазма содержит большое количество ядер и хлоропластов, как правило, лишенных пиреноидов. В цитоплазме имеются бесцветные лейкопласты (амилопласты), в которых откладывается крахмал. Вместо поперечных перегородок в талломе образуется внутренний скелет, сформированный особыми переплетающимися тяжами, по химическому составу сходными с внешними покровами таллома. Эти тяжи называют-
230