

5.1. Фракционный состав белков зерна злаковых культур
(% от общего количества белков)
|
Культуры |
Альбумины |
Глобулины |
Проламины |
Глютелины |
|
Пшеница |
10 |
15 |
30 |
35 |
|
Рожь |
25 |
20 |
20 |
25 |
|
Ячмень |
12 |
13 |
35 |
30 |
|
Овёс |
14 |
19 |
25 |
32 |
|
Кукуруза |
14 |
15 |
45 |
16 |
|
Просо |
12 |
10 |
55 |
13 |
|
Рис |
13 |
10 |
7 |
60 |
У мягкой и твердой пшеницы запасные белки образуют клейковину, количество и качество которой тесно коррелирует с хлебопекарными и мака-ронными свойствами зерна. Клейковина представляет собой сильно гидра-тированный белковый сгусток (студень), который выделяют из цельносмоло-того зерна при замешивании и отмывании водой получаемого из него теста. Такой белковый сгусток называют сырой клейковиной, в ней содержится в среднем 33–35 % сухого вещества. При высушивании сырой клейковины получают сухую клейковину. Отношение количества воды в сырой клейкови-не к количеству сухого вещества, выражаемое в процентах, называют показа-телем гидратации клейковины, который у пшеницы изменяется в пределах 170–250 %.
Сухая пшеничная клейковина на 80–90 % представлена запасными бел-ками – глиадинами (40–50 %) и глютенинами (35–40 %). Кроме запасных белков, в клейковине содержатся в небольшом количестве белки альбумино-глобулинового типа (3–6 %), крахмал (6–9 %), липиды (2–8 %), сахара (1–2 %), клетчатка (1–2 %), зольные вещества (0,5–2 %). Все химические компо-ненты клейковины объединяются в единый комплекс в результате возникно-вения водородных и дисульфидных связей, электростатических и гидрофоб-ных взаимодействий между группировками полипептидов, образующих мо-лекулы клейковинных белков.
Наличие большого количества нековалентных связей в структуре пше-ничной клейковины предопределяет её характерные физические свойства – упругость, эластичность, связность, растяжимость, способность к упругой де-формации. Фракции клейковинных белков глиадины (растворимы в 70%-ном растворе этанола) и глютенины (щёлочерастворимые белки) каждая в отдельности не формируют реологические свойства клейковины, они дей-ствуют совместно. Глиадины, образующие компактную пространственную структуру молекул, имеют внутримолекулярные дисульфидные (–S–S–) связи. Глютенины же характеризуются более рыхлой пространственной структурой молекул, способных образовывать межмолекулярные дисульфид-ные связи.
Реологические свойства клейковины зависят от соотношения в ней дисульфидных связей и восстановленных тиоловых групп (–SН). При увели-чении количества дисульфидных связей повышается упругость и связность клейковины, в результате чего она становится более крепкой. А если дисуль-фидные связи восстанавливаются с образованием НS-групп, упругость и связность клейковины понижаются и она утрачивает способность к упругой деформации.
Образование дисульфидных связей в клейковинных белках катализиру-ет фермент тиолоксидаза (1.8.3.2), который с участием кислорода способен окислять тиоловые группы и таким образом инициировать соединение в полимеры глютениновых субъединиц запасных белков в созреваюшем зерне. Расщепление дисульфидных связей катализирует фермент протеиндисуль-фидредуктаза (1.8.4.2), под действием которого происходит восстановление в белках дисульфидных связей с образованием тиоловых групп.
С участием указанных ферментов поддерживается определённое соот-ношение SS- и НS-групп в клейковинных белках, обеспечивая тот или иной уровень связности, упругости и эластичности клейковины. При увеличении в зерновках активности фермента тиолоксидазы и понижении активности про-теиндисульфидредуктазы отношение количества SS-связей к количеству HS-групп в клейковинных белках возрастает, вследствие чего улучшается качес-тво клейковины.
Клейковина активно участвует в формировании хлебопекарного теста, при взаимодействии с водой она набухает и образует упругую белковую структуру, соединяющую в единый комплекс все вещества теста. Выделяю-щийся в результате брожения сахаров углекислый газ растягивает клейко-вину, в результате чего тесто начинает увеличивать объём и подыматься, приобретая пористое строение. В тесте образуется большое количество по-ристых ячеек, наполненных СО2, и стенки этих пор в основном образованы белками клейковины. В процессе выпечки хлеба под воздействием высокой температуры пористая структура теста закрепляется вследствие денатурации белков, образуя характерный пористый хлебный мякиш.
Слишком крепкая клейковина формирует тугое тесто, которое с трудом поддаётся растяжению выделяющимся при брожении диоксидом углерода. Слабая клейковина не создаёт в тесте прочного белкового каркаса и СО2 в нём слабо удерживается. И в том, и другом случаях ухудшается пористость хлебного мякиша и уменьшается объём хлеба, что свидетельствует об ухудшении хлебопекарных свойств зерна.
Клейковина разжижается под действием протеолитических ферментов, подвергающих гидролизу запасные белки. Аналогичное действие оказывают протеазы, вводимые в зерновки клопом-черепашкой. Ослабление клейковины вызывает также фермент протеиндисульфидредуктаза, катализирующий рас-щепление дисульфидных связей с образованием восстановленных НS-групп.
Для характеристики качества зерна пшеницы обычно определяют сырую клейковину. Наряду с количеством проводится также оценка качества клейковины, главным образом по её эластичности и упругости, зависящих от состава и свойств запасных белков, образующих клейковину. Измерение упругости и эластичности клейковины выполняется с помощью специальных приборов, регистрирующих усилие, затрачиваемое на деформацию клейкови-ны. Результаты измерения выражают в виде показателя, называемого индек-сом деформации клейковины (ИДК). По этому показателю клейковину под-разделяют на 3 группы качества: I – хорошая, II – удовлетворительная, III – неудовлетворительная. Первая имеет ИДК в пределах 45–75, вторая – 80–100 и 40–20 , третья – 105–120 и 15–0.
По хлебопекарным качествам зерна мягкую пшеницу делят на три кате-гории: сильная пшеница, средняя (или ценная) и слабая. В зерне сильной пшеницы должно содержаться не менее 28 % сырой клейковины первой группы качества, в зерне ценной пшеницы – не менее 25 % сырой клейко-вины первой или второй группы качества. К слабой относится пшеница, в зерне которой содержится менее 25 % сырой клейковины или у которой клейковина очень низкого качества (третья группа).
Из сильной пшеницы получают муку, поглощающую большее количес-тво воды при замесе из неё теста, а получаемое из неё тесто устойчиво сохра-няет консистенцию, эластичность и форму, мало расплывается. Однако очень сильная пшеница образует тесто со слабой газоудерживающей способ-ностью, вследствие чего формируется пониженный объём хлеба. Такая пше-ница при добавлении к слабой пшенице улучшает её хлебопекарные свой-ства. Поэтому сорта сильной пшеницы называют сортами–улучшителями. Средняя пшеница способна давать хороший по качеству хлеб, но не может использоваться в качестве улучшителя слабой пшеницы. Слабая пшеница без добавления сильной пшеницы не способна давать удовлетворительный по качеству хлеб.
Хорошая по качеству твёрдая пшеница имеет крепкую клейковину, ко-торая очень упругая и плохо растяжимая. Поэтому она даёт хлеб с низким объёмом и плотным мякишем. При смешивании твёрдой пшеницы с мягкой слабой пшеницей качество хлеба улучшается.
Кроме пшеницы, клейковина также содержится в зерновках ржи и ячменя, а также в семенах травянистых злаковых растений. Клейковина ржи и ячменя по сравнению с пшеничной труднее отмывается и имеет низкие реологические свойства. А клейковина семян некоторых травянистых злако-вых растений (отдельные разновидности пырея, эгилопса и др.) характе-ризуется хорошими физическими свойствами, в связи с чем данные генотипы злаковых растений используются в селекции пшеницы как генетические источники для улучшения хлебопекарных свойств зерна.
В зерновках злаковых растений, кроме белков, содержатся в неболь-шом количестве и другие азотистые соединения: свободные аминокислоты и их амиды, пептиды, азотистые основания и нуклеотиды, нуклеиновые кисло-ты и др. На их долю приходится 5–10 % от общего количества азотистых веществ зерна. В наибольшем количестве они находятся в зародыше и алейроновом слое.
Белковые фракции зерна различаются по аминокислотному составу и в том числе по содержанию незаменимых аминокислот, от которых зависит биологическая ценность белков (табл. 1.4). Наиболее высокую биологичес-кую ценность имеют водорастворимые белки – альбумины, в их составе валин, лизин, треонин, триптофан содержатся почти в оптимальных соотно-шениях, однако в них понижено содержание метионина и фенилаланина и наблюдается небольшой дефицит по лейцину и изолейцину.
Солерастворимые белки зерна – глобулины также характеризуются довольно хорошо сбалансированным аминокислотным составом, хотя содер-жание некоторых незаменимых аминокислот у них по сравнению с альбу-минами понижено (метионин и лизин).
Щелочерастворимые белки – глютелины у ряда злаковых культур (овёс, ячмень, сорго) по содержанию незаменимых аминокислот близки к глобулинам, а у других (пшеница, рожь, кукуруза, просо) характеризуются довольно сильным дефицитом лизина, триптофана и метионина. Наиболее полноценны по содержанию незаменимых аминокислот глютелины риса – оризенины, в них отмечается лишь некоторый дефицит по метионину и лизину.
Самую низкую биологическую ценность имеют спирторастворимые белки – проламины, которые накапливаются только в семенах злаковых растений. Они отличаются очень низким содержанием таких незаменимых аминокислот, как лизин, триптофан, метионин, и высокой концентрацией глутаминовой кислоты, лейцина и пролина, на долю которых приходится 35–50 % от массы этих белков.
В связи с тем, что в зерне злаковых растений содержится много прола-минов и глютелинов, имеющих значительный дефицит лизина, изолейцина, метионина, треонина, валина, триптофана (табл. 1.3) суммарный белок их зерновок имеет довольно низкую биологическую ценность: белок зерна ку-курузы – 52–58 %, пшеницы, ячменя, проса – 60–70 %, ржи и овса – 70–75 %.
При изучении белковых фракций зерна современными физико-хими-ческими методами (различные способы хроматографии, электрофорез, изо-электрофокусирование и др.) установлено, что они состоят из большого набора индивидуальных белков, которые различаются по химическим свой-ствам и выполняют конкретные биологические функции.
В ходе разделения водо- и солерастворимых белков зерна злаковых культур (альбумины и глобулины) методом электрофореза в полиакриламид-ном геле можно идентифицировать на электрофореграммах до 30 белковых компонентов, различающихся по аминокислотному составу и молекулярным массам. Причём каждый генотип злаковых растений характеризуется опреде-лённым набором электрофоретических компонентов водо- и солераство-римых белков, в связи с чем по электрофореграммам этих белков проводится проверка генетической чистоты существующих сортов и идентификация новых генотипов зерновых культур.
Как показали результаты исследований, спирторастворимая фракция белков зерна пшеницы, которая выделяется после экстракции альбуминов и глобулинов, включает два вида белков – глиадины и спирторастворимый глютенин. При увеличении доли спирторастворимого глютенина во фракции спирторастворимых белков снижается упругость клейковины и устойчивость хлебопекарного теста, что в целом является причиной ухудшения качества хлеба.
Основную часть спирторастворимой фракции зерна пшеницы представ-ляют глиадины, которые методом электрофореза в полиакриламидном геле в лактат-алюминиевой буферной системе (рН 3,1) разделяются на 13–17 компонентов. Молекулы глиадинов обладают повышенной способностью к образованию межмолекулярных комплексов и взаимодействию с другими химическими компонентами зерна, вследствие чего они активно участвуют в формировании реологических свойств клейковины. Указанными свойствами обладают также проламины ржи, ячменя и овса. Формирование межмолеку-лярных агрегатов глиадинов и их комплексов с другими веществами зерна происходит за счёт образования водородных и дисульфидных связей, а также гидрофобных взаимодействий.
Из глютелиновых белков наиболее хорошо изучены глютенины – щелочерастворимые белки зерна пшеницы, которые включают определённый набор прочно связанных между собой полипептидных субъединиц. Большая часть субъединиц глютенинов представлена глиадиноподобными полипепти-дами, а остальная часть включает полипептиды альбумино-глобулинового типа. Полипептиды глютенинов обладают очень высокой способностью к образованию комплексов с глиадиновыми белками и другими химическими компонентами зерна и таким образом влияют на формирование реологи-ческих свойств клейковины.
Синтез белков в созревающем зерне. Синтез и накопление белков в зерновках злаковых культур происходит в основном за счёт оттока азотистых веществ (главным образом аминокислот) из вегетативных органов, так как поглощение минерального азота корнями и использование его в биосинтети-ческих процессах после цветения сокращается (особенно у яровых культур). Наибольшее количество азотистых веществ поступает в формирующиеся зерновки из листьев, особенно верхнего яруса, меньше из колосковых чешуй и стеблей. После цветения в листьях, стеблях и колосковых чешуях активизи-руются процессы гидролиза высокомолекулярных веществ (полисахаридов, белков, липидов, нуклеиновых кислот) и усиливается отток образующихся низкомолекулярных продуктов в зерно.
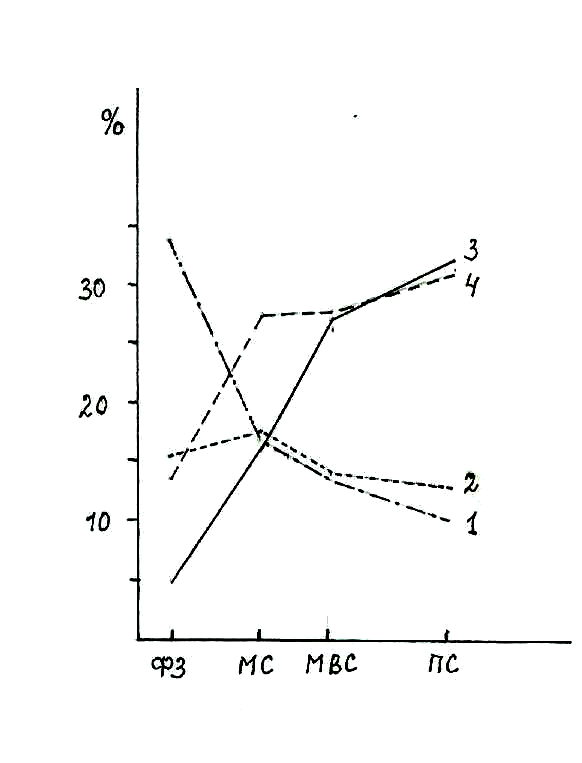
Рис. 5.1. Интенсивность синтеза различных групп белков в созревающем зерне пшеницы (поступление меченно-го 15N азота некорневой подкормки в белковые фракции зерна, %).
ФЗ – фаза формирования зерна; МС – молоч-ная спелость; МВС – молочно-восковая спе-лость; ПС – полная спелость; 1 – альбумины; 2 – глобулины; 3 – глиадины; 4 – глютенины.
В наших опытах с использованием меченного ¹⁵N азота показано, что на первых этапах формирования зерна в нём много содержится низкомолеку-лярных азотистых соединений, представленных в основном аминокислотами, а также легкорастворимых белков – альбуминов и глобулинов (рис. 5.1). В дальнейшем в ходе созревания зерновок концентрация в них небелковых азотистых веществ и белков альбумино-глобулинового типа постепенно снижается, но увеличивается накопление клейковинных белков – проламинов и глютелинов, при этом общее количество белков в зерне также увеличи-вается.
Поскольку в процессе созревания зерна относительное содержание альбуминов и глобулинов снижается, а количество проламинов и глютелинов увеличивается, в суммарном белке зерна наблюдаются соответствующие изменения концентрации аминокислот. Поскольку при созревании в зернов-ках увеличивается доля запасных белков с низким содержанием лизина, триптофана, метионина, треонина, изолейцина, то и в общем суммарном белке зерна также усиливается дефицит этих незаменимых аминокислот. Поэтому биологическая ценность суммарного белка в процессе созревания зерна снижается.
Такая закономерность в изменении аминокислотного состава суммар-ного белка особенно заметно проявляется в процессе созревания зерна пшеницы, кукурузы, ячменя, проса, сорго, у которых на долю проламинов приходится 30–50 % от общего количества белков в зерне.
У пшеницы по мере накопления запасных белков происходит форми-рование клейковинного комплекса зерна, в процессе созревания зерновок содержание клейковины повышается, улучшается её качество. В фазу мо-лочной спелости зерна клейковина имеет низкую гидратационную способ-ность и плохую связность, а к фазе полной спелости зерновок она приобре-тает характерные для неё реологические свойства. В ходе созревания зерна происходит постепенное уменьшение числа сульфгидрильных групп (–SН) в клейковине и увеличение количества дисульфидных связей вследствие сни-жения активности фермента протеиндисульфидредуктазы, катализирующего восстановление дисульфидных связей.