

2.3.3.2.2. Механизм сокращения
Способность мышечного волокна к сокращению обусловлена его ультраструктурой (рис. 17).
I-диск содержит только тонкие нити. Один конец такой нити прикреплён к Z–линии (рис. 16, 17), а другой – направлен к середине саркомера.
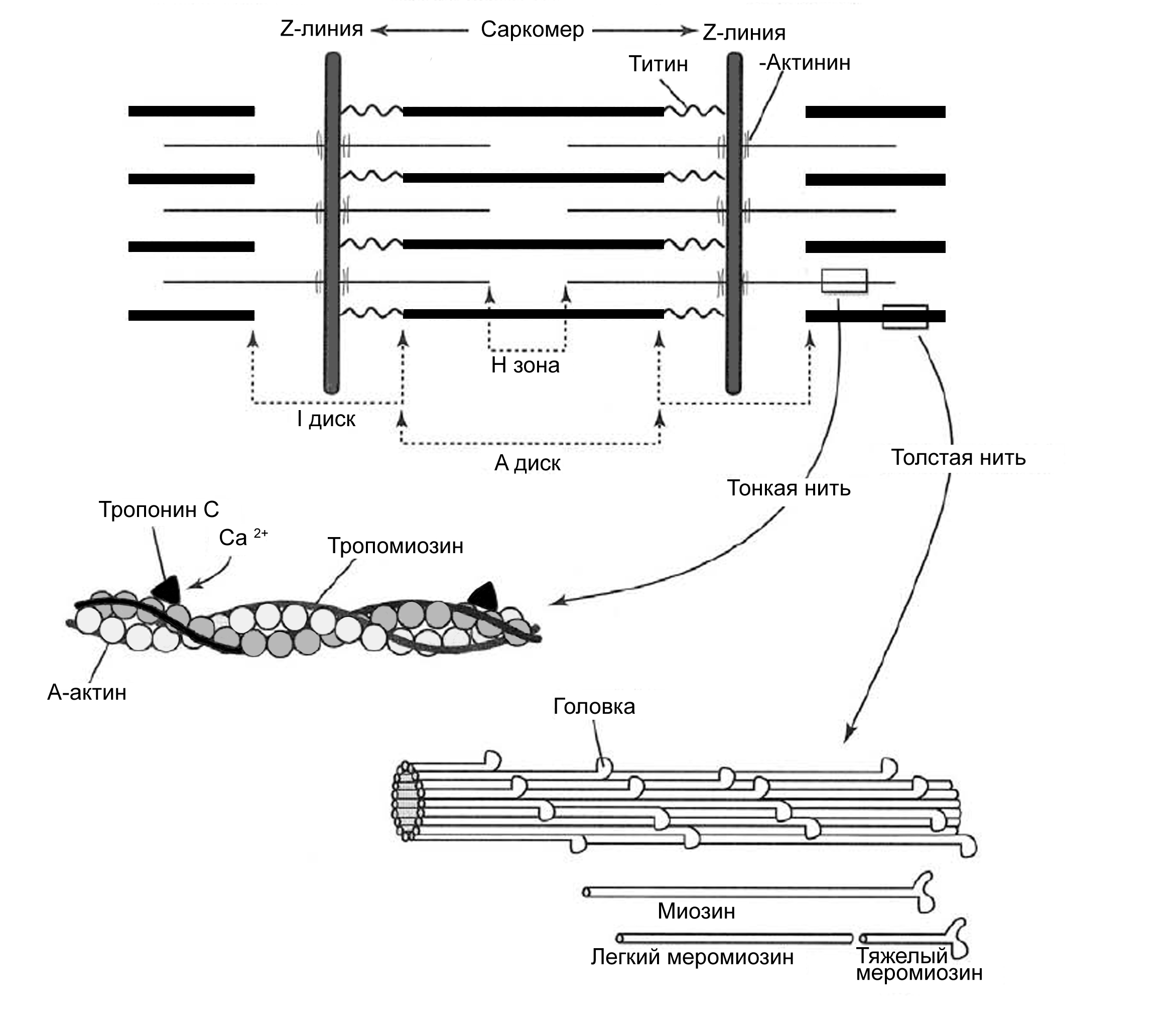
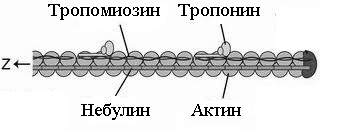
Рис. 17. Ультраструктура саркомера (по W. Ronald., 1990).
Толстые нити занимают центральную часть саркомера; их совокупность морфологически и обуславливает то, что называют А-диском. Тонкие нити лишь частично входят между толстыми. Следовательно, в саркомере имеется участок, содержащий только толстые нити – так называемая Н-зона. В середине Н-зоны проходит М-линия (мезофрагма) (рис. 16).
Таким образом, I-диск входит в состав двух саркомеров; то есть каждый саркомер содержит один А-диск (тёмный) и две половины I-дисков (светлых).
Толстая нить образована несколькими белками. Прежде всего, каждая миозиновая нить состоит из 300-400 молекул миозина, а также С-белка. Половина молекул миозина обращена головками (см. ниже) к одному концу нити, а вторая – к другому. Свободные концы толстых нитей с Z-линией связывает титин.
Миозин. В молекуле миозина различают тяжёлый и лёгкий меромиозин. Тяжёлый имеет два субфрагмента (S): S1 образован глобулярными головками миозина, S2 – эластичным компонентом. Лёгкий меромиозин образует концевую часть хвостовой нити миозина длиной 100 нм.
Молекула миозина имеет два шарнирных участка, позволяющих ей изменять конформацию («сгибаться»). Один шарнирный участок находится в области соединения тяжёлого и лёгкого меромиозинов, другой – в области шейки молекулы миозина (S1-S2 соединение); при этом лёгкий меромиозин лежит в толще толстой нити, тогда как тяжёлый меромиозин (благодаря шарнирным участкам) выступает на её поверхность.
Титин (Tn) – наибольших размеров полипептид, входящий в состав толстых нитей: его молекулярная масса около 300 кД. Он работает наподобие «молекулярной пружины», обеспечивая структурную целостность миофибрилл во время сокращения. Один конец молекулы проникает в Z-линию и через -актинин связывается с молекулой титина соседнего саркомера. Другой конец молекулы приближается к М-линии и при помощи миомезина прикрепляется к свободному концу молекулы титина второй половины саркомера. В I-диске титин ассоциирован с тонкими нитями, а в А-диске белок связывается с толстыми нитями. Таким образом, контактируя в области Z-линии, молекулы титина образуют непрерывную цепь.
С-белок стабилизирует структуру миозиновых нитей. Влияя на агрегацию молекул миозина, он обеспечивает одинаковый диаметр и стандартную длину толстых нитей.
Тонкая нить также образована рядом белков - актином, тропомиозином и тропонином.
Актин. Молекулы глобулярного актина (G-актин) полимеризуются и образуют фибриллярный актин (F-актин). Уложенные друг за другом молекулы F-актина в тонкой нити формируют две спирально закрученные цепочки.
Тропомиозин состоит из двух полипептидных цепей и имеет конфигурацию двойной спирали. Полярные молекулы тропомиозина длиной 40 нм укладываются «конец в конец» в желобке между описанными выше цепочками F-актина.
Тропонин – комплекс, образованный тремя глобулярными молекулами. Препятствует взаимодействию актина с миозином. Тропониновый комплекс прикреплён к молекулам тропомиозина, и потому он располагается с интервалами 40 нм.
Небулин – фибриллярный белок, ассоциированный с тонкими нитями.