

6. Измерение первичной продуктивности.
Идеальным способом измерения продуктивности было бы измерение потока энергии, идущего через систему, но пока сделать это трудно. В большинстве методов используются косвенные показатели – измеряется, например, количество произведённого вещества, количество использованного сырья или выделяющихся побочных продуктов. Каждый метод отражает какой-то один аспект сложного процесса автотрофно-гетеротрофного метаболизма. Упрощённое уравнение фотосинтеза:
hγ
6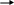 СО2
+ 6 Н2О
С6Н12О6
+ 6 О2
СО2
+ 6 Н2О
С6Н12О6
+ 6 О2
описывает общую реакцию образования углеводов из неорганических сырья за счёт световой энергии, действующей на хлорофилл. Поскольку в природе, как правило, продуцируется протоплазма, более полное уравнение процесса продукции должно выглядеть так:
1300000 кал hγ + 106 СО2 + 90 Н2О + 16 NО3 + 1 РО4 + минеральные элементы =
= 13000 кал потенциальной энергии, заключённой в 3258 г протоплазмы ( 106 С, 180 Н, 46 О, 16 N, 1 Р, 815 г зольных веществ) + 154 О2 + 1287000 кал рассеянной тепловой энергии (99%)
Это уравнение основано на соотношении элементов в протоплазме и на содержании в ней энергии. Ясно, что теоретически возможно определить продуктивность, измерив количество любого из компонентов уравнения за интересующий нас период времени. Уравнения, подобные приведённому здесь, могут применяться для пересчёта (и взаимной перепроверки) измерений продуктивности в единицах энергии, расхода СО2, NО3, или РО, веса протоплазмы (или количества углерода, переведённого в форму пищи), расхода О2.
Одна из самых больших трудностей при измерении продуктивности конкретной экосистемы состоит в определении того, находится ли система в состоянии динамического равновесия. В такой системе поступление вещества и энергии равно по выходу. В большинстве случаев естественных систем продуктивность под воздействием внешних изменений переходит от одного временного состояния устойчивого равновесия к другому.
Некоторые методы
1. Определение по урожаю. В системах, в которых травоядные животные имеют небольшое значение, и стационарное состояние никогда не достигается, продуктивность можно определять по урожаю. Именно так обстоит дело с посевами однолетних с/х культур, поскольку в данном случае принимаются меры по защите продукции от насекомых и других вредителей, а продуктивность растёт от нуля в момент посева до максимума ко времени сбора урожая. Взвесить собранный урожай и определить его калорийность не сложно.
По одному урожаю можно примерно оценить чистую продукцию молодых лесов или лесопосадок. Метод оценки по урожаю неприменим там, где продуцируемая пища изымается по мере её образования, как это характерно для многих природных сообществ. Если консументы – крупные долгоживущие животные, то продуктивность сообщества можно определить по биомассе консументов, потребляющих пищу с постоянной скоростью. В этом случае первичная продуктивность оценивается по вторичной. Так, продуктивность пастбищ может выражаться числом голов скота, которые прокормятся на данном количестве гектаров.
Недостатки такого метода очевидны. Он не позволяет учесть пищу, использованную самими растениями и ассоциированными с ними микроорганизмами, т.е. измеряется только чистая продукция сообщества.
2. Измерение количества кислорода. Поскольку производство пищи пропорционально продукции О2, последняя может быть основой для определения продуктивности. В некоторых водных экосистемах можно измерять продукцию О2 методами суточной кривой. При этом на протяжении суток периодически измеряют содержание растворённого в воде О2. Продукцию О2 днём и его потребление ночью можно оценить по площади графика, заключённого под суточной кривой. Этот метод наиболее пригоден для экосистем проточных вод, например рек и эстуариев, и особенно полезен при изучении загрязнённых вод. Если существует заметная диффузия кислорода между водной массой и атмосферой, то это служит источником ошибки; впрочем, здесь легко ввести достаточно надёжный поправочный коэффициент, т.к. диффузия подчиняется хорошо известным физическим законам. Метод суточной кривой даёт валовую первичную продукцию, поскольку О2, потреблённый ночью, суммируется с кислородом, произведённым в дневное время суток (т.е. тем самым автоматически учитывается дыхание всего сообщества).
3. Определение двуокиси углерода. В наземных экосистемах удобнее измерять изменения количества СО2, а не О2. Физиологи растений равно используют такой показатель как потребление СО2, для измерения фотосинтеза в листьях или отдельных растениях, а агрономы и экологи много раз пробовали измерять продукцию в целых интактных сообществах, заключая их в прозрачные камеры. Большой колоколообразный сосуд, пластмассовый ящик или палатку помещают над сообществом, а через образовавшуюся камеру прогоняют воздух и измеряют концентрацию СО2 в нём на входе и на выходе.
4. рН-метод. В водных экосистемах рН воды зависит от содержания растворённого СО2, которое в свою очередь уменьшается в результате фотосинтеза и увеличивается за счёт дыхания. Но чтобы использовать рН в качестве показателя продуктивности, сначала надо построить калибровочную кривую для исследуемой воды, т.к.: 1) зависимость рН от содержания СО2 нелинейная и 2) степень изменения рН на единицу изменения содержания СО2 зависит от буферной ёмкости воды (т.е. расходование 1 ед. СО2 на фотосинтез вызовет в мягкой воде горной реки более сильное повышение рН, чем в хорошо забуференной морской воде).
5. Определение расхода сырья. Как видно из уравнения, приведённого ранее, продуктивность можно измерить не только по скорости образования веществ (пищи, протоплазмы, минеральных веществ), но и по скорости расхода минерального сырья. Правда, в системах с устойчивым равновесием расход может уравновешиваться поступлением из самих организмов или притоком извне, и тогда определить действительную скорость потребления организмами невозможно. Там, где такие компоненты, как N или Р, поступают не постоянно, а предположим, раз в год или через меньшие промежутки времени, прекрасным мерилом продуктивности за соответствующий период может служить скорость уменьшения концентрации этих веществ. Метод надо применять с осторожностью, т.к. сырьё может исчезать и безучастия организмов. Этим способом определяется чистая продукция всего общества.
6. Метод радиоактивных изотопов. Имея определённое количество «меченого» вещества, которое легко обнаружить по излучению, можно проследить скорость переноса даже в упомянутой выше системе с устойчивым равновесием, причём система к тому же почти не изменяется этим вмешательством.
7. Определение хлорофилла. Метод оценки растительной продукции основан на представлении о том, что интенсивность фотосинтеза определяется количеством хлорофилла. Морские водоросли ассимилируют максимум 3,7г углерода на 1 г хлорофилла в 1 ч. Общую продуктивность данного участка моря можно оценить, если известна концентрация хлорофилла на разной глубине и уменьшение освещённости с глубиной. Хотя этот хлорофилловый метод менее точен, чем методы, основанные на газообмене, он, тем не менее, позволяет довольно просто и быстро оценивать продуктивность океанов и озёр.