

Равенство Доннана.
При выводе уравнений,
описывающих распределение ионов между
клеткой и окружающей средой, выполняется
условие электронейтральности, равенство
суммарной концентрации анионов ( в
основном
и ионов макромолекул
![]() )
катионов
)
катионов
как внутри клетки
![]() ,
,
так и снаружи клетки
![]() .
.
Здесь - число отрицательных зарядов на каждой белковой молекуле.
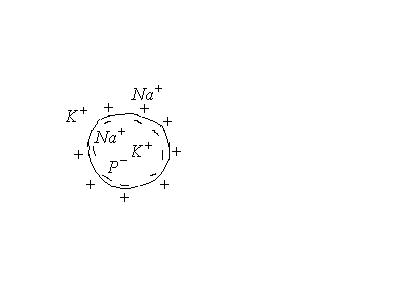
В межклеточной жидкости содержание катионов значительно выше, чем ионов макромолекул . Поэтому
![]() - равенство Доннана
- равенство Доннана
Потенциал покоя
Экспериментально установлено, что цитоплазма в состоянии покоя имеет отрицательный потенциал, а окружающая среда- положительный.
Действительно, в первом приближении
В клетке
![]() в 20-40 раз.
в 20-40 раз.
Снаружи
![]() в 10 раз.
в 10 раз.
Это неравномерное
распределение концентраций обусловлено
![]() насосом, при котором при переносе
насосом, при котором при переносе
![]() переносится
переносится
![]() .
Кроме того, в клетке имеются анионы
макромолекул
(белков, аминокислот, и др.).
.
Кроме того, в клетке имеются анионы
макромолекул
(белков, аминокислот, и др.).
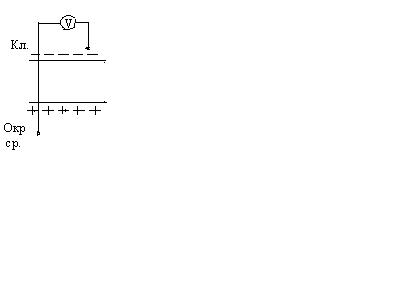
Ионы внутри клетки не связаны с другими ионами и могут диффундировать туда, где их мало, т.е. в окружающую среду. Т.е. в состоянии покоя клетка проницаема только для ионов . Анионы не могут проникать через мембрану и остаются на внутренней поверхности мембраны. Т.о. мембрана снаружи зарядится положительно, а внутри – отрицательно.
Между внутренней и внешней поверхностями мембраны возникает разность потенциалов мембранной природы. Эта разность потенциалов между клеткой и окружающей средой, измеренная в состоянии физиологического покоя, называется потенциалом покоя.
Если принять, в первом приближении, что потенциал покоя определяется только диффузией ионов , то величина потенциала покоя определяется как
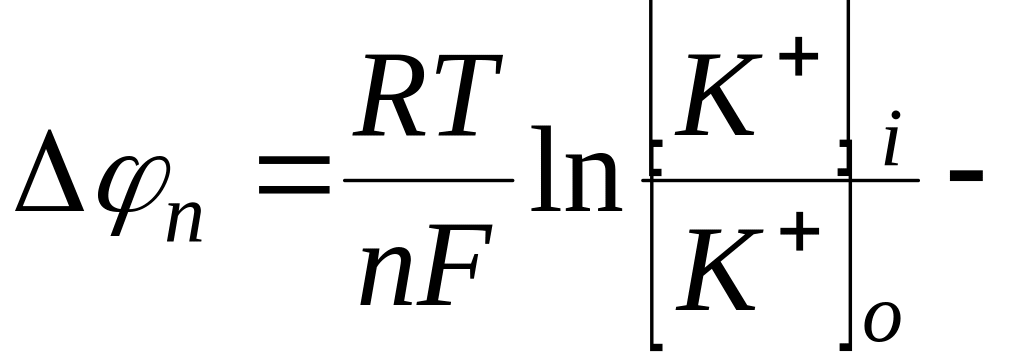 уравнение
Нернста.
уравнение
Нернста.
![]() - активная
концентрация ионов
внутри клетки,
- активная
концентрация ионов
внутри клетки,
![]() - активная
концентрация ионов
снаружи.
- активная
концентрация ионов
снаружи.
Если
=
,
то
![]() клетка мертва.
клетка мертва.
Но
![]() ,
поэтому
,
поэтому
![]() .
Например, для аксона гигантского кальмара
.
Например, для аксона гигантского кальмара
![]()
![]() ,
а это подтверждает, что в основе
возникновения потенциала действия
лежит перенос ионов.
,
а это подтверждает, что в основе
возникновения потенциала действия
лежит перенос ионов.
В реальности в
состоянии покоя мембрана проницаема
не только для ионов
,
но и для ионов
и
.
Например, для аксона гигантского кальмара
экспериментально установлено, что
![]() .
Основной вклад в потенциал покоя вносят
ионы
и
.
Ионов
переносится очень мало. Поэтому на
внешней поверхности
.
Основной вклад в потенциал покоя вносят
ионы
и
.
Ионов
переносится очень мало. Поэтому на
внешней поверхности
сосредоточивается
положительный заряд,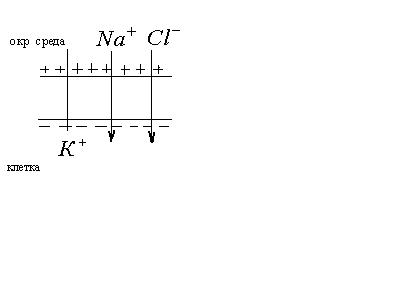 а на внутренней – отрицательный.
Потенциал покоя определяется тремя
диффузионными потоками и вычисляется
(с учетом равенства Доннана) по формуле
а на внутренней – отрицательный.
Потенциал покоя определяется тремя
диффузионными потоками и вычисляется
(с учетом равенства Доннана) по формуле
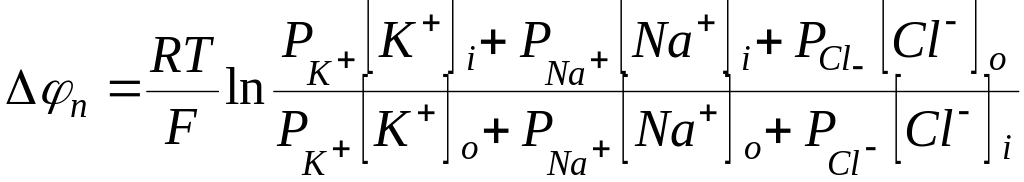 .
.
Кроме простой диффузии ионов и может идти обменная диффузия, но, как мы отмечали, потоки их равны и обменная диффузия не влияет на мембранный потенциал.
Потенциал действия.
Все клетки возбудимых тканей (нервная, мышечная, железистая) под действием различных раздражителей достаточной силы способны переходить в возбужденное состояние. Обязательным признаком возбуждения является изменение электрического состояния мембраны.
Опыт показывает, что возбужденный участок становится
электроотрицательным по отношению к
невозбужденному участку. Следовательно, на возбужденном участке происходит перераспределение ионов. При возбуждении это перераспределение кратковременно и концентрации восстанавливаются после снятия возбуждения, а разность потенциалов становится равной исходной, т.е. потенциалу покоя. Для аксона кальмара обнаружена такая зависимость изменения потенциала при возбуждении от времени
- потенциал покоя; мембранный потенциал при возбуждении;
- общее изменение разности потенциалов.
Общее изменение разности потенциалов между клеткой и окружающей средой, происходящее при пороговом и сверхпороговом возбуждении клетки, называется потенциалом действия.
Механизм возникновения потенциала действия. В 1938 году Круэл и Картис показали, что сопротивление аксона кальмара в состоянии покоя 1000 Ом/см2, а при возбуждении 25 Ом/см2, т.е. уменьшается в 40 раз. При этом сопротивление цитоплазмы не изменяется. Следовательно, уменьшение сопротивления мембраны обусловлено только её проницаемости для ионов, т.к. именно они являются переносчиками электричества в мембранах и клетках.
Хаджкин, Хаксли и Катц показали, что при возбуждении проницаемость мембраны увеличивается только для ионов , причем в 500 раз. Это приводит к увеличению диффузии ионов из окружающей среды в клетку (по концентрационному градиенту), что приводит к изменению потенциала мембраны. В первые моменты возбуждения интенсивность потока ионов
из клетки остается такой же, как и до возбуждения. Поэтому поток ионов вызывает исчезновение избыточного отрицательного потенциала на внутренней поверхности мембраны. Эта фаза называется деполяризацией и длится короткое время. Затем начинается другая фаза – реполяризация., заключающаяся в следующем. Диффузия ионов внутрь клетки нарушает равновесие концентраций в клетке. В связи с этим повышается проницаемость мембраны для ионов , начинается диффузия ионов из клетки в окру-жающую среду. Поток ионов из клетки приводит к уменьшению проницаемости для ионов . В результате происходит реполяризация мембраны и восстановление потенциала покоя. Проницаемость мембраны для ионов и падает до исходной величины. Фаза реполяризации длится дольше фазы деполяризации, поэтому и кривая более пологая.
В некоторых случаях регистрируется так называемый следовой потенциал, как на данном рисунке. Он вызван тем, что после окончания возбуждения проницаемость мембраны для ионов и остпется повышенной.
Т.о. формирование потенциала действия обусловлено двумя потоками через мембрану: поток внутрь клетки приводит к перезарядке мембраны, а противоположный поток обусловливает восстановление потенциала. Потоки эти приблизительно равны по величине, но сдвинуты по времени. Благодаря этому сдвигу во времени и возможно появление потенциала действия.