

Сокращение
Сокращение мышечного волокна (рис. 18) происходит при поступлении возбуждения по аксонам двигательных нейронов к нервно-мышечным синапсам в виде нервных импульсов (потенциал действия нервных волокон). Это - непрямое сокращение (оно опосредовано нервно-мышечной синаптической передачей). Прямое же сокращение (например, мышечные подёргивания, или фибрилляции) – патология.
Сокращение мышечного волокна происходит при взаимодействии толстых и тонких нитей. В покое взаимодействие тонких и толстых нитей невозможно, так как миозинсвязывающие участки молекул актина заблокированы тропомиозином. При поступлении импульса (как уже отмечалось) из Т-системы по каналам саркоплазматической сети в сарколемму поступают ионы кальция. При высокой их концентрации эти ионы связываются с комплексом TnC и вызывают конформационные изменения тропомиозина. Это, в свою очередь, приводит к разблокированию миозинсвязывающих участков молекул актина, благодаря чему головки миозина начинают поочерёдно присоединяться к молекулам актина тонкой нити и изменяют свою конформацию. Таким образом, создаётся тянущее усилие, и тонкие нити начинают скользить между толстыми.
При этом головка миозина совершает около 5 циклов в секунду; когда одни головки миозина толстой нити производят тянущие усилия, другие в это время свободны и готовы вступить в очередной цикл. Следующие друг за другом «гребковые» движения (молекула миозина сгибается) стягивают тонкие нити к центру саркомера. Таким образом, скользящие тонкие нити тянут за собой Z-линии, вызывая укорочение саркомера.
В процесс сокращения практически одновременно вовлечены все саркомеры мышечного волокна, за счёт чего и происходит его укорочение (сокращение).
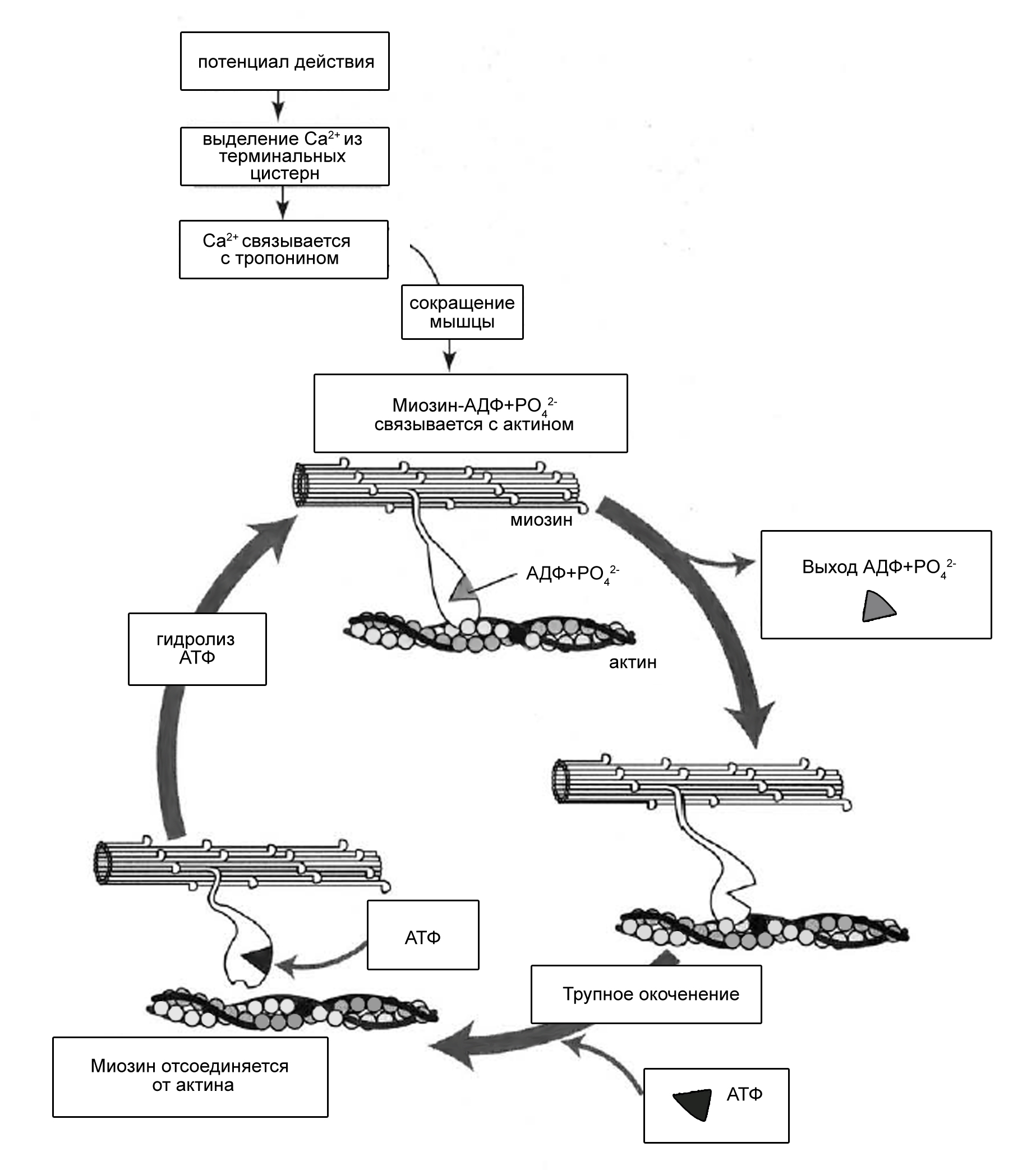
Рис. 18. Механизм сокращения мышечного волокна (по W. Ronald, 1990).
Расслабление
Как уже указывалось
выше, Са![]() -АТФаза
саркоплазматической сети закачивает
ионы кальция из саркоплазмы в цистерны
ретикулума, где они связываются
кальсевестрином. При понижении
концентрации ионов кальция до определённого
уровня Са
-тропомиозин
закрывает, блокирует миозинсвязывающие
участки и вновь препятствует их
взаимодействию с миозином.
-АТФаза
саркоплазматической сети закачивает
ионы кальция из саркоплазмы в цистерны
ретикулума, где они связываются
кальсевестрином. При понижении
концентрации ионов кальция до определённого
уровня Са
-тропомиозин
закрывает, блокирует миозинсвязывающие
участки и вновь препятствует их
взаимодействию с миозином.
После смерти организма, когда содержание АТФ в мышечных волокнах снижается вследствие прекращения её синтеза, головки миозина оказываются устойчиво прикреплёнными к тонкой нити. Это состояние трупного окоченения продолжается до тех пор, пока не начнётся разрушение волокон.
2.3.3.2.3. Гистогенез и регенерация поперечнополосатой мышечной ткани
В онтогенезе скелетная мускулатура образуется из миотомов, где формируются клетки-предшественники вытянутой формы – миобласты. На этой стадии часть миобластов обособляется, давая начало камбиальным клеткам – сателлитам.
Многократно поделившись, миобласты последовательно сливаются друг с другом, образуя следующую стадию – миотубы. В них начинается сборка миофибрилл – сократительных структур с характерной поперечной исчерченностью, что заканчивается формированием зрелого поперечнополосатого мышечного волокна.
Регенерация. После повреждения разрушенные фрагменты волокон фагоцитируются. В сохранившейся части волокна начинаются процессы его восстановления; на концах таких волокон образуются «почки», растущие навстречу друг другу. В свою очередь сателлиты, плотно прилежащие к поверхности симпласта, активно делятся; образовавшиеся клетки либо включаются в состав мышечных почек, либо сливаются между собой, образуя новый симпласт (то есть происходит процесс, аналогичный гистогенезу этой ткани).
Часто при восстановлении мышечной ткани образуется соединительнотканный рубец.
Описанная структура поперечно-полосатого мышечного волокна в различных группах животных весьма варьирует. Например, у членистоногих длина саркомера даже у одного животного в различных мышцах различна и может составлять от 0,5 до 15 мкм. В широких пределах изменяется соотношение тонких и толстых протофибрилл: в наиболее быстрых мышцах оно составляет 3:1 (почти как у позвоночных), но может быть и 5:1 и даже 6:1. В таких мышцах обычно нарушается правильное расположение тонких нитей вокруг толстых.
В мышцах членистоногих большую вариабельность имеет и строение Z-линий, вплоть до того, что в некоторых они практически не выделяются.
Весьма своеобразным вариантом поперечнополосатых мышечных тканей являются асинхронные летательные мышцы насекомых. По интенсивности работы они представляют собой наиболее совершенную механохимическую систему: в ответ на один нервный импульс эти мышцы могут совершать до 300 сокращений в секунду - при этом длина саркомера уменьшается лишь на 2%, а не на 20%, как в обычных мышцах. Есть у этих мышц и ещё ряд своеобразных особенностей.
У высших позвоночных животных различают три типа мышечных волокон: быстросокращающиеся красные, быстросокращающиеся белые, медленносокращающиеся промежуточные (как отмечалось выше, окраска мышц обусловлена миоглобином). Соответственно, в зависимости от преобладания того или иного типа волокон, скелетные мышцы называют «красными» и «белыми», либо «быстрыми» и «медленными». Доля тех или иных волокон различна в зависимости от степени «загруженности» мышц – например, у спортсменов, занимающихся разными видами спорта, этот показатель различен.
Различные типы волокон отличаются как структурно, так и функционально (табл. 9):
Таблица 9