

§ 6.4. Внутриводоемный круговорот пероксида
ВОДОРОДА И РЕДОКО-СОСТОЯНИЕ
ВОДНОЙ СРЕДЫ
Одним из измеряемых в гидрохимии параметров природной воды является ее окислительно-восстановительный (редокс-) потенциал Eh-Эта термодинамическая по своей сути характеристика отражает суммарное относительное содержание в водной среде окислительно-восстановительных реагентов. Поскольку в воде обычно присутствует растворенный кислород, потенциал платинового электрода, помещенного в воду, имеет положительный заряд по сравнению с водородным электродом сравнения. Иначе говоря, среда окислительная. Отрицательное значение редокс-потенциала имеет место лишь в анаэробной среде, когда вместо Ог в воде присутствует, например, H2S. Эту ситуацию можно охарактеризовать как восстановительную или даже сверхвосстановительную.
207
Реальная природная вода находится не в термодинамическом, а в динамическом окислительно-восстановительном равновесии в том смысле, что окислительные и восстановительные реагенты в ней постоянно образуются и взаимопревращаются. При этом можно различать по крайней мере две редокс-подсистемы, основанные на участии в окислительно-восстановительных процессах молекулярного кислорода (медленно устанавливающееся равновесие) и пероксида водорода (быс-трореагирующая подсистема).
Измеряемое значение Eh при этом определяется процессами, протекающими главным образом в "медленной" подсистеме.
При рассмотрении окислительно-восстановительных процессов в природных водах присутствие в аэробной водной среде пероксида водорода следует рассматривать как само собой разумеющееся, поскольку Н2С>2 — промежуточный продукт круговорота кислорода в окружающей среде. Образование ЩОг на пути восстановления Ог до воды практически неизбежно, за исключением частных случаев.
Первое сообщение об обнаружении Н2О2 в природной воде (в водах Мексиканского залива) было сделано американскими гидробиологами Бааленом и Марлером в 1966 г. Концентрация пероксида водорода была незначительной, порядка 10"7 М, и сообщение Баалена и Марле-ра не привлекло широкого внимания ученых. Исключение составил советский ученый В.Е. Синельников. Он изучал явление хемилюминес-ценции в природных водах и применил метод хемилюминесценции для измерения малых концентраций Н2О2.
В конце 60-х — начале 70-х годов В.Е. Синельников провел большое число измерений содержания Н2О2 в различных природных водах нашей страны, главным образом на Волге. Везде и всюду пероксид обнаруживался, хотя концентрация его изменялась в широких пределах, достигая в ряде случаев 10"5 М. На основе большой совокупности полученных данных В.Е. Синельников приходит к выводу, что пероксид водорода является неотъемлемым компонентом природной водной среды.
С начала 80-х годов интерес к природному пероксиду водорода резко возрос, появилось множество публикаций по измерению Н2Ог в объектах водной среды. Обнаружение Н202 в природных водах уже не кажется удивительным, скорей удивительно, если его нет.
Много Н2Ог в атмосферной влаге. В дожде содержание Н2Ог дости гает 10~4 М, в снеге — 10"5 М. Исследования, проведенные в Гренлан дии, показали, что Н2О2 содержится даже в ископаемых льдах, возраст которых исчисляется десятками тысяч лет. Это позволило сделать вывод, что в атмосферной влаге в аэробной среде пероксид водорода всегда присутствовал. Вместе с осадками пероксид водорода поступал 208 /
и на сушу, и в поверхностные воды суши. Пероксид водорода образуется в природных водах и в результате внутриводоемных процессов.
Рассмотрим более подробно процессы, приводящие к образованию Н2О2 в природных водах.
Упомянутое уже поступление из атмосферы в виде "готового" Н202. Измерения, проведенные американскими исследователями в Атлантическом и Тихом океанах, показали, что после дождя содержание Н2О2 в поверхностных водах океана резко возрастает, но в течение нескольких часов уменьшается до некоторого стационарного уровня, обусловленного внутриводоемными пррцессами. Этот уровень претерпевает суточные колебания, что указывает на роль солнечного излучения в образовании Н2О2.
Основную роль в образовании Н202 в природных водах играет солнечный свет, под действием которого образуется супероксидный радикал:
Н02 + О' ~» 02 + НО" (1)
Исследования с применением фермента супероксиддисмутазы и акцепторов Or-радикала показывают, что супероксидный радикал является предшественником большей части Н202, образующегося в природных водах при воздействии солнечного УФ-света.
3. Возможно двухэлектронное восстановление 02 до Н202 в редокс- каталитическйх процессах по реакции
02
+ DH2
fo2*/011*
» D
+
н202 (2)
Конечно же в редокс-каталитических процессах пероксид водорода может образовываться и через одноэлектронные стадии восстановления 02 и тогда его предшественником вновь будет супероксидный радикал.
Отмеченное В.Е. Синельйиковым увеличение содержания Н202 в зоне выброса неочищенных городских сточных вод указывает на то, что вклад каталитических процессов' окисления легкоокисляемых органических веществ в образование Н202 может быть существенным.
4. Биологическая эмиссия служит наиболее существенным источни ком Н202 при наличии в воде микроводорослей. Известны многие виды водорослей, выделяющие Н202 во внешнюю среду в процессе фотосинтеза. Практически все микроводоросли продуцируют Н202 при действии УФ-составляющей солнечного излучения. Происходит это вследствие протекания внутриклеточных фотохимических реакций с участием светочувствительных водорастворимых компонентов клетки.
Образующиеся под действием УФ-света радикалы 02 рекомбинируют
209
в клетке (с участием супероксиддисмутазы) и избыточный пероксид водорода выводится во внешнюю среду.
Помимо микроводорослей известны некоторые бактерии, выделяющие Н2О2 во внешнюю среду в процессе жизнедеятельности.
5. К образованию Н202 могут приводить некоторые гидроперокси-ды, образующиеся в фотохимических и каталитических процессах при окислении органических веществ синглетным кислородом или свободными радикалами:
![]()
(3)
Содержание ROOH в природных водах обычно примерно в 10 раз меньше, чем Н2О2.
В общем виде скорость образования Н202 в природных водах складывается из абиотического и биотического вкладов и может быть представлена в виде
wf — Wfti + аз/>2Д
где tu/д
Л77[Нг02],М
скорость абиотических реакций образования Н202; аз/>2 —
эффективная константа скорости образования Н202 в биотическом канале; В — биомасса микроводорослей.
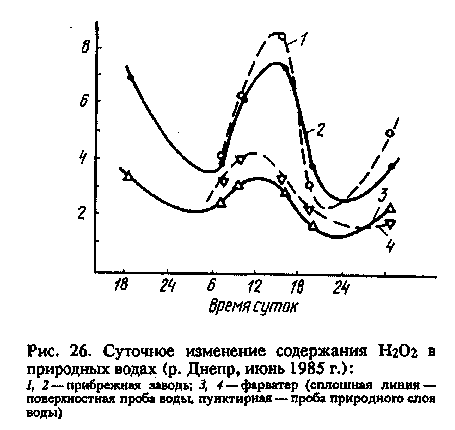
Содержание пероксида водорода в водной среде изменяется в течение суток (рис. 26), что свидетельствует о протекании наряду с образованием также и процессов разложения Н202. Размах суточных колебаний и средний уровень содержания Н202 в водной среде зависят от сезона (температура), погодных факторов (солнечный свет) и гидрологических особенностей водоема (проточность, эффективность перемешивания слоев воды и т.д.).
Если добавить пероксид водорода в природную воду, то скорость его распада зависит от многих факторов, прежде всего от присутствия микроводорослей и от интенсивности солнечного излучения. В филь-> 210
трованной воде скорость разложения Н202, как правило, крайне низка. На солнечном же свету в нефильтрованных пробах она даже ниже, чем в темноте. Последнее обстоятельство связано с тем, что скорость образования Н2О2 при фотолизе водорослей выше, чем скорость ее фотохимического разрушения.
В фильтрованных пробах основной процесс разложения Н202 — его каталитический распад на содержащихся в воде ионах меди и железа, а также фотохимическое разложение.
10sw0,M4
В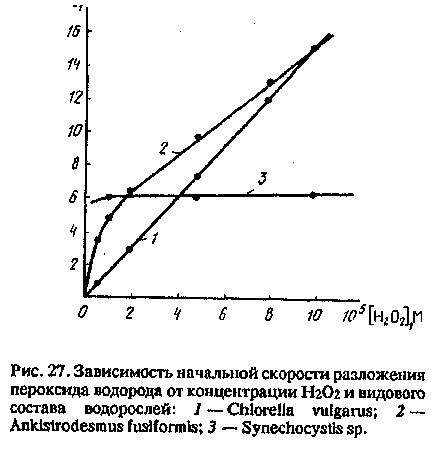 нефильтрованных пробах скорость
разложения Н2О2 пропорциональна
биомассе микроводорослей и сопутствующих
им бактерий. Характерная особенность
кинетики разложения Н2О2 в природных
водах — осуществление двух процессов:
первого и нулевого порядка по
концентрации Н2О2 (рис. 27).
нефильтрованных пробах скорость
разложения Н2О2 пропорциональна
биомассе микроводорослей и сопутствующих
им бактерий. Характерная особенность
кинетики разложения Н2О2 в природных
водах — осуществление двух процессов:
первого и нулевого порядка по
концентрации Н2О2 (рис. 27).
Реакция первого порядка дает больший вклад при высоких концентрациях Н2О2. Наличие двух процессов связано с действием различных ферментативных систем микроорганизмов. Реакция первого порядка обусловлена вкладом каталазного процесса разложения Н2О2 до кислорода и воды. Механизм реакции нулевого порядка более сложен. Хотя в разрушении Н2О2 непосредственно участвует пероксидаза, но субстраты пероксидазного окисления поставляются дегидрогеназными реакциями, в ходе которых образуются вещества восстановительной природы — доноры Н. В стационарных условиях скорость реакции разложения Н2О2 нулевого порядка определяется скоростью образования восстановителей, взаимодействующих с Н2О2 с участием пероксидазы в качестве катализатора.
Скорость биологических реакций разрушения И2Ог как первого, так и второго порядка пропорциональна биомассе водорослей:
Щ = («41 + 8е(£(2[Н202])Д
причем
211
![]()
эе^д В = щ ~ аэгД
![]()
где аэ^ь md,2, «г — эффективные константы скорости соответствующих процессов; wr — скорость образования восстановительных эквивалентов — перехватчиков Н202 в дегидрогеназных реакциях, пропорциональная биомассе водорослей.
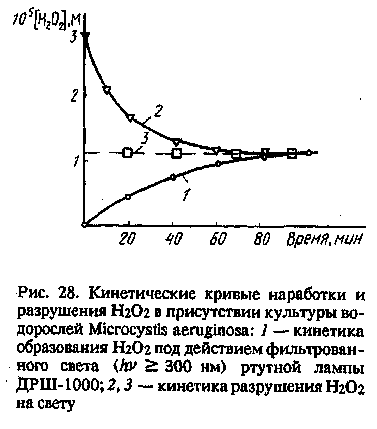
Если природную воду или культуру водорослей одновременно подвергать воздействию солнечного УФ-излучения и добавлять туда разные количества Н2О2, то по прошествии некоторого времени независимо от исходной концентрации Н2О2 в воде установится примерно одна и та же стационарная концентрация Н2О2 (рис. 28). Может быть подобрана такая исходная концентрация Н202, при которой она не изменяется во времени. Значение этой стационарной концентрации сильно зависит от вида водорослей или состава природного альгоценоза. В случае водорослевых культур стационарная концентрация достигает 10"5 — 3 • 10~5 М и, по-видимому, соответствует содержанию внутриклеточной Н2О2 при воздействии УФ-излучения.
Стационарная концентрация Н202 в водной среде в естественных условиях определяется из равенства скоростей ее образования и разрушения. Приравнивая выражения для этих скоростей, легко получить
![]()
или, опуская абиотический вклад в образование Н2О2 (при аэ^д w эег)> находим
![]()
Наличие разности двух величин в этих выражениях указывает на возможность реализации двух принципиально различных состояний природной водной среды (если разность положительна, то поток Н2О2 превышает поток восстановителей; если отрицательна — поток восста-
212
новителей превышает поток реакционноспособных окислительных эквивалентов).
Следует подчеркнуть, что восстановительные эквиваленты, продуцируемые биотой, могут более эффективно окисляться пероксидом водорода, чем растворенным кислородом. В противном случае эти восстановители служат дополнительным источником Н2О2 в водной среде:
DH2 + 02 -+ D + Н202 (4)
DH2 + Н202 -► D + 2Н20 (б)
Например, аскорбиновая кислота каталитически окисляется кислородом воздуха более эффективно, чем пероксидом водорода. Соответственно при окислении ее кислородом в среде накапливается Н2Ог. Напротив, гидрохинон более эффективно окисляется пероксидом водорода. При каталитическом окислении гидрохинона кислородом воздуха Н2Ог в среде не накапливается.
Поскольку стационарное содержание Н202 в природных водах применено на два порядка ниже, чем растворенного 02, восстановители, регулирующие содержание в водной среде Н202, должны окисляться кислородом по крайней мере на два порядка менее эффективно, чем пероксидом водорода. При низкой природной концентрации катализаторов, в частности ионов меди, такой относительный выигрыш в эффективности процессов окисления Н202 по сравнению с 02 может быть объяснен тем, что восстановление Ог, как правило, требует участия двух восстановленных ионов металла, тогда как восстановление Н202 — одного.
При преобладании потока восстановителей над потоком Н202 стационарная концентрация DH2 будет превышать концентрацию перок-сида водорода. В этом случае при определении окислительно-восстановительных эквивалентов "быстрой" подсистемы методом титрования вместо окислителя (Н2О2) будет обнаруживаться восстановитель (DH2). Наиболее наглядно качественное различие между двумя редокс-состоя-ниями водной среды следует из сравнения графиков титрования проб воды пероксидом водорода (рис. 29).
В том случае, если отрезок отсекается на оси ординат (в воде присутствует Н202 природного происхождения), речь идет о нормальном окислительном состоянии среды. Если же отрезок отсекается на оси абсцисс, то можно говорить о квазивосстановительном состоянии аэробной водной среды. Приставка "квази" указывает на временный характер такого состояния, поскольку при наличии в воде растворенного кислорода рано или поздно (в замкнутой "медленной" подсистеме) произойдет окисление всех восстановительных эквивалентов.
213


214
Видно, что в темповых условиях в природной воде при конкурентной борьбе сине-зеленых водорослей за среду обитания идет процесс накопления веществ-восстановителей, титруемых Н202- Скорость образования DH2 зависит от экологической обстановки и изменяется в широких пределах.
Природа восстановителей пока не ясна, но известно, что среди них присутствуют серосодержащие (тиольные) соединения.
Особенностью тиольных соединений является взаимодействие их с ионами меди, сопровождающееся восстановлением Си2+ в Си+ и образованием прочных, малореакционношособных комплексов с Си*. Так,. механизм окисления глутатиона (GSH) — одного из природных восстановителей, регулирующих внутриклеточные редокс-процессы, — ионами меди в присутствии 02 и Н2О2, — описывается следующими реакциями:
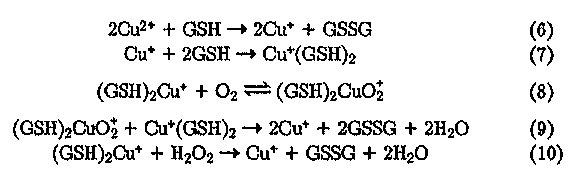
Из реакций (6)—(10) видно, что при низких концентрациях меди скорость окисления глутатиона кислородом воздуха может быть гораздо меньше, чем пероксидом водорода. Фактически глутатион инактивирует медь, выводя ее из каталитического редокс-цикла. Соответственно при формировании в природной воде квазивосстановительных условий значительно снижается способность водной среды к самоочищению (см. гл. 7).
Переход от нормальных окислительных условий к квазивосстановительным сопровождается значительным изменением качественных характеристик природной водной среды. В то же время стандартные гидрохимические характеристики ее, такие, как рН, Eh, 02, ХПК, БПК и другие, не претерпевают сколь-нибудь заметных изменений.
Происходит такой переход обычно в водоемах, подверженных цветению сине-зеленых водорослей, незадолго до их массовой вегетации при повышении температуры до 18—20° С, при безветренной погоде. Образующийся в фотохимических процессах пероксид водорода в квазивосстановительной среде расходуется на "титрование" восстановительных эквивалентов, выделяемых сине-зелеными водорослями.
Наиболее существенное следствие изменения редокс-состояния водной среды заключается в появлении в воде не известных ранее
215
факторов токсичности. В качестве биотестов могут быть взяты инфузории, дафнии, личинки рыб на самых ранних стадиях их развития. Так, инфузории Tetrahymena pyriformis, клетки которых делятся каждые 5 ч, при острой токсичности погибают в течение часа, а при хронической выживают, но заметно снижается их способность к размножению. Если к остротоксичной пробе воды добавлять малыми порциями Н202, то токсичность ее постепенно снижается и после оттитровывания восстановительных эквивалентов среда становится биологически полноценной. Тем самым можно говорить о факторе редоке-токсичности водной среды.
Если в качестве тестовых организмов рассматривать личинки рыб, то они реагируют на редокс-состояние не сразу, а накапливают токсический эффект, который наиболее остро проявляется при переходе личинок на жаберное дыхание. Фактически личинки задыхаются при наличии в среде достаточного количества растворенного кислорода. Другими словами, при формировании в воде, в которой находятся личинки, квазивосстановительных условий они теряют способность усваивать кислород. Если в эту воду ввести ионы меди (в количестве, превышающем количество восстановителя), то личинки нормально развиваются. Снимается эффект токсичности и при периодических (для непрерывного поддержания окислительных условий среды) добавках Н2(>2 в исходных концентрациях порядка 10"4 М. Эти наблюдения показывают, что в квазивосстановительной среде возникает дефицит по биологически доступной меди, участвующей в формировании активных центров дыхательных ферментов.
И дефицит Н2О2 в среде сам по себе также может быть токсичен для личинок рыб. Участие экзогенного (наружного) пероксида водорода в процессах жизнедеятельности личинок было установлено на некоторых видах рыб в аквариумных опытах.
В течение многих лет невольным участником крупномасштабного токсикологического "эксперимента" являлся Волгоградский осетровый рыбоводный завод (ВОРЗ), построенный в целях компенсации ущерба, причиненного строительством Волгоградской ГЭС естественному воспроизводству этой ценной породы рыб. Известно, что 90% мировых запасов осетровых сосредоточены в Волго-Каспийском регионе и их естественные нерестилища расположены в основном на Нижней и Средней Волге. Стада осетровых (осетр, белуга, севрюга) собираются под Волгоградской плотиной и вынуждены нереститься в малопригодных для этого местах. Часть производителей (самцов и самок) отлавливают и с помощью специальных рыбоводных методик получают созревшую икру, искусственно ее осеменяют и разводят сначала личинок, а потом молодь рыб в рыбоводных прудах. 216
Многолетние наблюдения показали, что выживаемость личинок в течение рыбоводного сезона сильно меняется в зависимости от редокс-состояния водной среды. Усредненная качественная картина сезонного изменения выхода личинок, перешедших на смешанное питание, представлена на рис. 31, из которого видно, что, пока в воде есть Н202 (среда окислительная), выживаемость личинок близка к 100%. В период неустойчивого редокс-состояния внешней среды (днем Нг02 появляется, к утру исчезает) выход личинок резко снижается. Когда среда полностью переходит в квазивосстановительное состояние (НгСЬ не обнаруживается и днем), -то выход личинок в этих условиях близок к нулю.
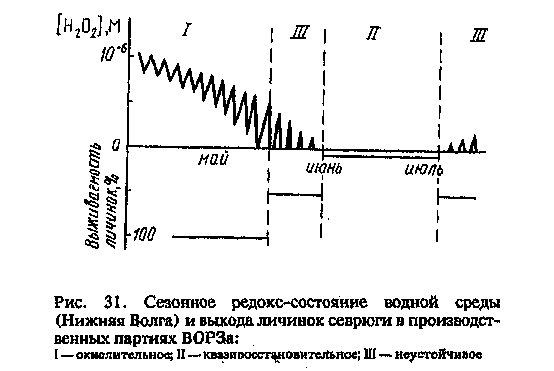
Сроки наступления и продолжительность периодов неустойчивого и квазивосстановительного состояния среды могут меняться от сезона к сезону, но качественный характер сохраняется.
Считается, что непосредственной причиной (или ближайшим следствием) гибели личинок является нарушение у них жироуглеводного обмена: жир не перерабатывается в гликоген, и личинка погибает, когда исчерпываются материнские запасы гликогена. Не исключено, что личинки, выжившие в условиях неустойчивого редокс-состояния среды, несут в себе отпечаток от этого периода (несбалансированность жироуглеводного обмена) всю жизнь. И этот дефект может проявиться в следующие периоды жизни особи, когда единственным источником энергии становятся жировые запасы, например в преднерестовый период. Не исключено, что отмечающееся в последние годы расслоение мышечной ткани (жировыми прослойками) у производителей
217
осетровых — следствие перенесенного ими в личиночной стадии дефицита Н2О2 и биологически доступных ионов меди.
До начала 70-х годов пероксид водорода в волжской воде всегда обнаруживался. В начале 80-х вместо Н2Ог в воде стали находить вещества-восстановители, титруемые пероксидом водорода. Если принять, что массовая гибель личинок связана с формированием в воде квазивосстановительных условий, то состояние волжской водной среды начало "менять знак" в 1976 г., когда это явление было впервые отмечено на ВОРЗе. Именно в этом году были введены в строй сооружения биологической очистки наиболее крупных городов в бассейне р. Волги.
Оказалось, что в заводских условиях можно успешно бороться с этим новым токсичным фактором, добавляя в воду, в которой выращиваются личинки, пероксид водорода и ионы меди (в нетоксичных концентрациях). При таком искусственном поддержании окислительного состояния среды личинки нормально развиваются, переходят на активное питание и в выростных прудах быстро набирают вес. Жиро-углеводный обмен у таких личинок протекает нормально — жир переходит в гликоген.
Гораздо хуже обстоит дело с естественными водоемами. Ведь те же заводские личинки, помещенные в волжскую воду, погибали. Перокси-да водорода для поддержания окислительного состояния всей реки не напастись. Здесь нужны другие подходы: как вернуть внутриводоем-ным редокс-процессам необходимый для нормального функционирования водной экосистемы баланс?
Не только осетровые, но и другие виды рыб, нерестовый период которых приходится на период формирования квазивосстановительных условий среды, обречены на резкое сокращение воспроизводства. Во всяком случае на заводах по искусственному разведению растительноядных рыб также участились случаи повышенных отходов — гибели производственных партий личинок при формировании в воде'квазивосстановительных условий среды.
Возникновение этих условий предшествует бурному развитию сине-зеленых водорослей. По-видимому, вещества-восстановители представляют собой одну из разновидностей альготоксинов (см. § 6.2). Одним из следствий изменения редокс-состояния среды является развитие в квазивосстановительной среде патогенной микрофлоры. Так, с распространением возбудителей чумки в период "цветения" сине-зеленых водорослей связана гибель балтийских и байкальских тюленей. Видимо, и другие проявления ухудшения санитарно-токсикологической обстановки в водах, подверженных "цветению" сине-зеленых водорослей, связаны с расбалансировкой внутриводоемных редокс-процессов. 218
Токсикация водной среды в отношении аэробных водных организмов на личиночной стадии их развития или микроорганизмов с интенсивным водообменом с внешней средой может быть связана с действием одного или нескольких факторов редокс-токсичности.
Непосредственное действие восстановителя как токсиканта (блокирование активных центров окислительно-восстановительных ферментов, нарушение внутриклеточного редокс-равновесия и др.).
Перевод меди в биологически недоступную форму и, как следствие, возникновение дефектных ферментных систем (апоферментов).
Отсутствие в среде Н2Ог как экзогенного окислителя, участвующего при нормальных условиях в пероксидазном окислении эндогенных субстратов на ранних стадиях развития водных организмов до формирования ими собственных систем пероксидазного окисления.
Снижение способности водной среды к самоочищению и связанное с этим возможное накопление токсических веществ (фенолов, аминов, нитратов и др.), которые были бы окислены при наличии в среде цероксида водорода.
б. Создание благоприятных условий для возникновения и размножения патогенной микрофлоры,