

Глава 9
ТОКСИЧЕСКОЕ ДЕЙСТВИЕ ЗАГРЯЗНЯЮЩИХ ВЕЩЕСТВ
Одной из важнейших задач по охране водных объектов является проблема нормирования качества воды и нагрузки на водоем как по отдельным загрязняющим веществам (так называемым приоритетным загрязнителям), так и по их совокупности. Нынешняя концепция, основанная на использовании системы предельно допустимых концентраций, недостаточна, поскольку она ориентирована главным образом на санитарно-гигиенические требования и нацелена на охрану здоровья человека, а не на защиту "здоровья" экосистем.
Для установления экологических нормативов необходимы новые подходы к оценке токсического воздействия отдельных ЗВ и их совокупности на уровне не только отдельных особей или популяций, но и всей экосистемы. Задача эта сложная, поскольку токсические эффекты существенно зависят от таких показателей, как температура, содержание в воде кислорода, рН, качественный состав РОВ (наличие в воде лигандов, редокс-агентов, антиоксидантов и т.д.). Кроме того, попадающие в природную среду вещества, трансформируясь, могут значительно менять свои токсикологические характеристики, что также затрудняет нормирование. В свете этого для более адекватного подхода к оценке токсикологических эффектов необходимо развитие экологической токсикологии на базе изучения молекулярных механизмов воздействия различных ЗВ на физиологические процессы в клетке и в экосистеме.
Микроорганизмы эволюционируют, поддерживая низкие внутриклеточные концентрации токсических веществ. Адаптация к токсическим веществам происходит двумя путями. Первый путь, обеспечивающий устойчивость к токсинам, по-видимому, носит наследственный характер и ведет свое начало от организмов, которые появились на Земле в экстремальных природных условиях. Встречаются же популяции бактерий и водорослей, устойчивых к высоким концентрациям ионов металлов, обитающих при низких значениях рН и.при высоких температурах. Горячие источники, вулканические озера и глубокие кратеры на дне морей обеспечивают такие экстремальные условия, которые были характерны Для Земли миллиарды лет назад. Другой
293
путь связан с созданием биохимических механизмов, обеспечивающих устойчивость к токсическим факторам. Так, у некоторых бактерий устойчивость развивалась в результате синтеза молекул экстрахромосомной ДНК (плазмидов). Синтезируемые некоторыми, организмами внутриклеточные полимеры связывают и удаляют из клетки ионы металлов (Cd, Cu, Ni, Co). Ферментативное окисление или восстановление токсичных загрязняющих веществ, попадающих внутрь клетки, может переводить их в менее токсичные формы и т.д.
Исследование биохимических основ токсичности и устойчивости животных организмов к действию загрязняющих веществ очень важно для адекватной оценки возможных экологических последствий тех или иных антропогенных воздействий. Для этого необходимо изучение принципов транспорта и превращения ЗВ в живых клетках, чего нельзя понять без знания основных процессов жизнедеятельности.
§ 9.1. Общие сведения о структуре и функции
КЛЕТКИ
Говоря о токсичности водной среды, необходимо различать несколько аспектов: токсичность в отношении человека и теплокровных животных, отдельных видов растений и насекомых (гербициды, инсектициды), низших представителей трофической цепи (бактерий, микроводорослей) и токсинов в отношении экосистемы в целом. Иначе говоря, токсичность тех или иных факторов окружающей среды должна рассматриваться на уровне организмов, популяций и экосистем.
В любом случае необходимо знать как бы элементарные механизмы токсических эффектов. При этом не обойтись без изучения воздействия различных токсикантов и физических факторов на внутриклеточные процессы и на межклеточную информационную систему.
Клетка — основная единица живого объекта, очаг жизни. Она отделена от внешней среды плазматической мембраной, ограничивающей и регулирующей диффузию ионов и молекул из внешней среды во внутреннюю и обратно. Мембраны не только отделяют клетку от внешней среды, но и создают архитектуру клетки. В некоторых случаях мембраны составляют до 80% общей массы сухих компонентов клетки. Увидеть клеточные мембраны в разрезе возможно лишь с помощью электронного микроскопа.
Функции плазматической мембраны — регулирование переноса веществ в клетку и из нее, обеспечение взаимодействия с соседями, проведение электрических сигналов в нервных и мышечных волокнах. Плазматическая мембрана клетки неоднородна. Одни фрагменты 294
имеют гладкую поверхность, другие образуют характерные выпячивания и складки.
Непосредственно к плазматической мембране примыкают структуры, составляющие скелет клетки (цитоскелет). Эти структуры образованы двумя типами элементов: тонкими нитями из актиноподобного белка (актин — основной белок тонких мышечных волокон), пронизывающими всю цитоплазму, и микротрубочками диаметром около 25 А, образуемыми белком турбулином. Основная функция цитоскелета — опорная, поддержание жесткой конструкциии. В то же время "трубочки" могут участвовать и в переносе веществ.
Действие любого токсиканта на внутриклеточные процессы так или иначе связано с проникновением его внутрь клетки. В основе биохимии окружающей среды лежит понимание процессов, определяющих транспорт элементов через клеточные мембраны. Физические и химические параметры, такие, как рН, температура, окислительно-восстановительные условия и природа загрязняющего вещества, сильно влияют на механизм транспорта и токсичность загрязнителя. Кроме того, клетки, являясь сложной химико-биологической системой открытого типа, обмениваются с внешней средой веществами, участвующими в поддержании процессов жизнедеятельности, и информационными экстраметаболитами. Итак, первым барьером на пути токсиканта выступает плазматическая мембрана.
Способность клетки к обмену веществами с окружающей средой приводит к возникновению разности их концентраций во внутри- и внеклеточном пространстве. Разность концентраций может поддерживаться лишь при условии, что мембрана обладает различной проницаемостью для разных веществ.
В состав мембраны входят в основном фосфолипиды и белки. Соотношения этих компонентов изменяются от 2,5:1 до 1:2,5 в зависимости от функции мембраны.
Фосфолипиды представляют собой эфиры трехатомного спирта — глицерина и жирных кислот с длинными углеводородными радикалами:
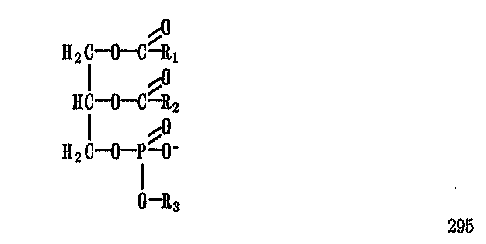
Ацильные остатки жирных кислот могут сильно отличаться по длине цепи, ее разветвленности и по числу двойных связей.
Жирнокислотные остатки липидных молекул мембран клеток животных представляют собой неразветвленные цепи с четным числом атомов углерода (14—-24). Структура наиболее распространенных мембранных фосфолипидов может быть представлена формулой
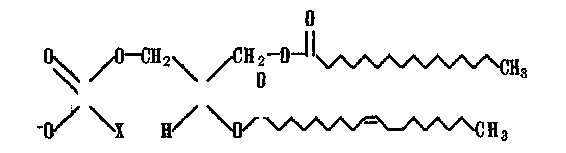
Р С II
Фосфолипиды плохо растворяются и в воде, и в неполярной фазе, поскольку имеют в своем состайе полярную (голова) и гидрофобную (хвост) группы.
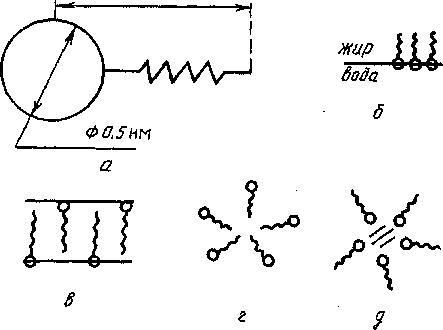
Молекулы таких соединений образуют истинные растворы, находясь в мономерном состояний, липгь при чрезвычайно низких концентрациях. В водном окружении они располагаются так, чтобы гидрофобные неполярные остатки были изолированы от контакта с водой. В силу этой молекулярной особенности фосфолипиды располагаются на поверхности раздела вода — неполярная фаза либо образуют двойной слой или глобулы, мицеллы (рис. 43).
Знм
296
Рис. 43. Схематическое изображение молекул фосфолипидов (а) и их расположение на границе раздела жир-вода (б), в бислойной мембране (в), при образовании неполярной (г) и полярной (д) мицелл
Строение липидных агрегатов зависит от многих факторов: химической природы фосфолипида, его концентрации, степени гидрофиль-ности среды, температуры, наличия или отсутствия в среде поверхностно-активных веществ и др. Устойчивые бислойные структуры образуются обычно при повышении объемной доли липидов до 50%.
Все фосфолипиды обладают сходными свойствами. Они способны образовывать комплексы с холестерином и полярными группами других молекул, таких, как белки.
Холестерин — вещество стероидной природы — обязательный компонент плазматических мембран клеток жийотных, где он присутствует и в свободном состоянии, и в виде эфиров с жирными- кислотами. Холестерин влияет на расположение фосфолипидов в мембране, а его наличие в большом количестве в некоторых мембранах имеет физиологическое значение.
При смешении фосфолипидов с холестерином образуется бимолекулярный липидный слой толщиной 4—10 нм. При этом холестерин, являющийся слабополярным липидом (имеет одну полярную группу —• ОН), легко "протыкает" мембрану.
Белки адсорбируются на поверхности мембраны, образуя липопро-теиновый слой, легко проницаемый для гидрофобных веществ и малопроницаемый для ионов и полярных молекул. В зависимости от того, требует ли перенос вещества через мембрану затраты дополнительной энергии, эти процессы могут быть разделены на пассивные (при наличии градиента концентрации или электрохимического потенциала) и активные (требующие затраты энергии).
Жирорастворимые нейтральные молекулы проходят через мембрану путем простой диффузии через липидные мембраны в направлении градиента концентрации
где S — площадь поверхности мембраны, через которую осуществляется транспорт вещества Р; Л с — градиент концентрации, d — толщина мембраны; К — коэффициент диффузии, зависящий от коэффициента распределения вода — неполярная фаза (растворимости Р в липиде), молекулярной массы вещества Р, его пространственной структуры.
В сдучае гидрофильных молекул малых размеров (г < 4 А) - Н20, ,Н202, метанол, мочевина и др. — возможна фильтрация через мелкие поры (каналы) в мембране. Эти каналы появляются и исчезают в силу динамического характера липидопротеинового слоя.
Для отдельных клеток обмен с внешней средой может осуществ-
10*-235 297
ляться также путем молекулярного пиноцитоза и э к о ц и -т о з а — образование вакуоли или микрокапли внеклеточной жидкости.
Этот статистический динамический процесс, обусловленный отсутствием жесткого каркаса мембраны, служит важным механизмом транспорта макромолекулярных веществ внутрь клетки и выделения их во внешнюю среду.
Аналогичный процесс переноса твердых частиц через мембрану называют фагоцитозом.
Специфический (селективный) характер имеет широко используемый в природе так называемый активный транспорт, когда вещество Р переносится через мембрану с помощью специальных ферментов-переносчиков (трансфераз) против градиента концентрации или электрохимического потенциала. Этот процесс требует затраты энергии в виде АТФ. Благодаря активному транспорту в клетке концентрируются вещества, находящиеся во внешней среде в низких концентрациях. Он поддерживает и контролирует оптимальный состав внутриклеточной" среды.
Активный перенос осуществляется, в частности, в случае ионов Na+, К+ и С1", при этом действует своего рода "ионный насос", откачивающий ионы Na+ из клетки. Так, активность нервных и мышечных клеток связана с изменением ионной проницаемости мембран в отношении Na+, K+. Проницаемость мембраны управляется с помощью ацетилхолина, проникающего в мембрану мышечной клетки в месте стыковки с нервным окончанием.
Итак, вещество Р по одному из перечисленных каналов проникло внутрь клетки. Здесь происходит его межфазовое распределение в соответствии с коэффициентом распределения вода — неполярная фаза (см. § 7.2). Полярные соединения будут сосредоточены в основном в цитоплазме, неполярные — в липидных внутриклеточных мембранах.
Вещества, обладающие комплексообразующей способностью, т.е. содержащие донорные или акцепторные группы (орбитали), будут образовывать донорно-акцепторные и комплексные соединения. Внутриклеточным комплексообразователем в плазме является белок альбумин, точнее его N-концевая аминокислота — аспарагин. Обычно эта кислота занята эндогенными субстратами, например жирными кислотами. Связывание субстратов с альбумином происходит конкурентно.
Кроме альбумина высокой комплексообразующей способностью обладают нуклеиновые кислоты. Комплексообразование с ними может угнетать зависящий от ДНК синтез РНК.
Некоторые вещества обладают лигандными свойствами и могут конкурентно с эндогенными субстратами образовывать комплексные 298
соединения с ионами металлов в активных центрах металлоферментов. С другой стороны, ионы тяжелых металлов могут образовывать координационные соединения с клеточными лигандами, в качестве которых выступают главным образом белки.
Говоря о клетке, необходимо иметь в виду ее пространственную неоднородность. Схематическое изображение "внутренности" клетки представлено на рис. 44.
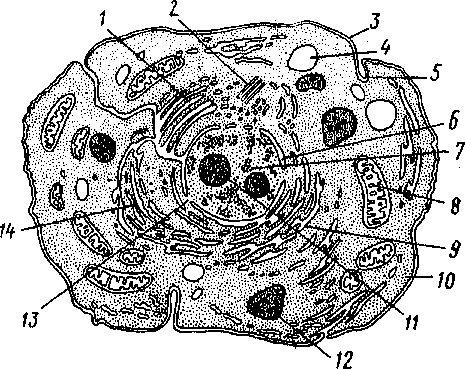
Рис. 44. Основные органеллы клетки:
1 — комплекс Гольджи; 2 — микросома; 3 — плазматическая мембрана; 4 — вакуоль;5—пиноцитарный пузырек; б— ядерная мембрана; /—ядро;* — митохондрии; 9-—эндоплазматический ретикулум; 10—цитоплазма; И — ядрышко; 12 — лизосома; 13 — поры в ядерной мембране; 14 — рибосомы
Внутриклеточные частицы, находящиеся в цитоплазме, называют органеллами. Они отличаются друг от друга структурой и функциональным назначением. В цитоплазме расположено клеточное ядро, не всегда выделенное морфологически. Например, у самых простых клеток — бактерий и сине-зеленых водорослей — нет явцо выраженного ядра, но в цитоплазме содержатся скопления веществ, характерные для ядер клетки более сложных организмов. Ядро служит местом хранения и воспроизведения большей части наследственного материала клетки и представляет собой частицу, ограниченную двойной мембраной. Ядерная мембрана имеет множество каналов — крупных пор (50 нм), через которые в цитоплазму могут проходить самые различные вещества, но задерживается хромосомный материал клетки.
Во всех клетках, за исключением эритроцитов и бактерий, цито-
299
плазма представляет собой не однородную массу, а сложную систему мембран, образуемых так называемым эндоплазматическим ретикулу-мом. Он состоит из обширной сети трубочек, пузырьков и цистерн, крайне разнообразных с морфологической точки зрения. Система эндоплазматического ретикулума очень лабильна в структурном и функциональном отношении. Степень ее развития изменяется не только в клетках разного типа, но и на разных стадиях развития или физиологического состояния.
На мембранах эндоплазматического (шероховатого) ретикулума, как и на мембране ядра, со стороны цитоплазмы прикреплены рибосомы, представляющие собой "гранулы" РНК, в которых идет синтез белка (ферментов). Часть рибосом свободно плавает в основном веществе цитоплазмы. Синтезированный белок проходит через мембрану эндоплазматического ретикулума и по трубочкам переносится в определенные места клетки.
Гладкие мембраны эндоплазматического ретикулума (комплекс Гольджи) образуют субклеточные частицы особого типа — лизосомы. Лизосомы представляют собой пузырьки, окруженные одиночной мембраной и содержащие гидролические ферменты для расщепления практически любого компонента клетки. Мембрана этих органелл несет большую функциональную нагрузку — при ее повреждении содержимое лизосом прорывается в клетку и полностью растворяет ее. В состав эндоплазматического ретикулума входят также микросомы, представляющие "очистные сооружения" клетки.
Во всех клетках аэробных организмов содержался митохондрии — своего рода "энергетические станции". В них протекают основные метаболические процессы образования и накопления энергии, необходимой для функционирования клетки. В зависимости от типа клетки в ней может находиться от нескольких десятков до нескольких тысяч митохондриальных частиц.
Митохондрии образованы двумя мембранами: внешней гладкой и внутренней, образующей многочисленные складки, которые проникают глубоко внутрь органеллы. Вещества с небольшой молекулярной массой легко проникают через наружную мембрану митохондрий. Проникновение веществ во внутримитохондриальное пространство, а также выход из него строго контролируются системами внутренней мембраны.
Митохондрии представляют собой структурный комплекс ферментов цикла трикарбоновых кислот (цикл Кребса). Это место образования АТФ. Энергия для окислительного фосфорилирования АДФ берется в результате постепенного окисления субстратов — доноров Н в дыхательной цепи: Сахаров, липидов, аминокислот. Ферменты, обес-300
печивающие перенос восстановительных эквивалентов, являются частью внутренней мембраны митохондрий.
Кроме того, в клетке имеются пероксисомы, в которых осуществляются процессы пероксидазного окисления эндогенных субстратов, и липосомы — центры синтеза липидов мембран.
В растворимой фракции клеток (цитозоле) находятся многие окси-дазы (алкогольоксидаза, альдегидоксидаза, ксантиноксидаза), некоторые дегидрогеназы (алгольдегидрогеназа), а также восстановительные ферменты (редуктазы), катализирующие восстановление соединений с
двойной связью, дисульфидов, сульфоксидов ((CHa^SO-*—*->(CH3)2S),
N-оксидов
((CH3)3NO
l2Kh
(CH8)8N),
а также дегидроксилирование
ароматических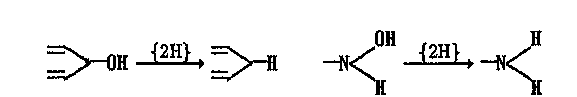 соединений
и гидроксиламинов:
соединений
и гидроксиламинов:
Рассмотрим схему обмена веществ в клетке. В качестве "пищи" (органического топлива) клетка использует жиры, углеводы и белки.
Наиболее энергоемкими хранителями энергетических запасов являются жиры (у животных) и масла (у растений). Это сложные эфиры длинноцепочечных карбоновых кислот и 1,2,3-триоксипропана (глицерина) - так называемые триглицериды (ТГ):
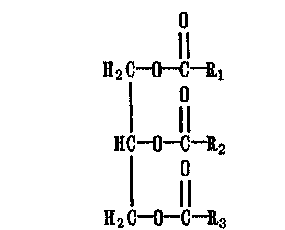
где Ri, R2, R3 содержат от 3 до 21 атома С, в том числе и одну или несколько ненасыщенных связей в уио-конфигурации.
В результате гидролиза триглицеридов в воде образуются глицерин и карбоновые кислоты жирного ряда с четным числом атомов С — от 12 до 22.
Растительные масла содержат преимущественно ненасыщенные, легкоокисляемые жирные кислоты, животные жиры — насыщенные жирные кислоты (ЖК).
301
Гидролиз ТГ осуществляется с помощью ферментов липаз:
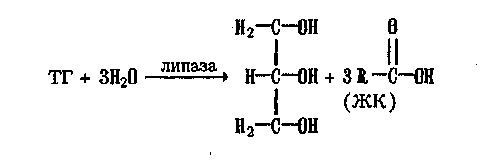
Образующиеся при этом ЖК дегидрируются, гидрагируются и расщепляются, происходит их так называемое /2-окисление, в осуществлении которого активное участие принимает кофермент А (КоА):
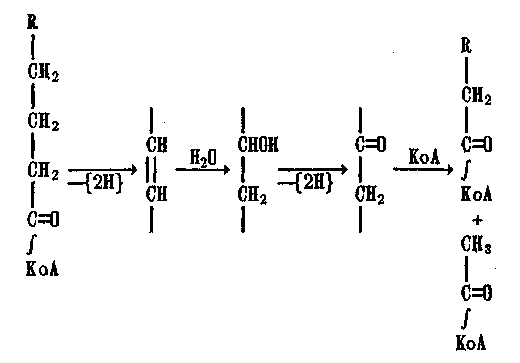
Процесс расщепления ЖК продолжается с накоплением ацетил-КоА.
Глицерин дегидрируется, изомеризуется и декарбоксилируется с образованием уксусного альдегида:
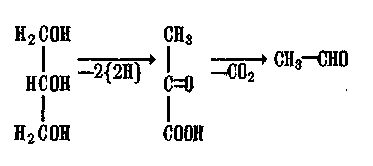
Углеводы (общая формула Са^НгО)^) легко отдают запасенную в них энергию на оперативные нужды клетки. Это полимеры глюкозы (крахмал, гликоген, целлюлоза), различающиеся по способу полимеризации а-, /?-форм глюкозы. Гликоген формирует энергетический
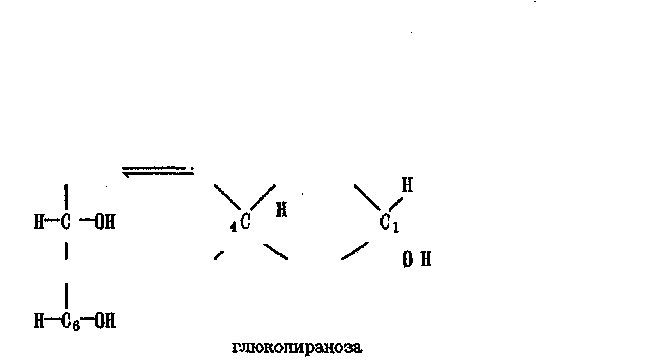
резерв клетки, целлюлоза является строительным материалом растений. Древесина на 50% состоит из целлюлозы, а хлопок — почти на 100%. Ферменты, катализирущие гидролиз гликогена, целлюлозу не расщепляют. В результате гидролиза крахмала и гликогена образуется глюкоза, находящаяся в растворе в линейной либо циклической форме. Циклическая форма (глюкопираноза) полимеризуется за счет образования молекул воды при комбинации ОН-групп при Ci и Cj:
О
II H-Ci
I
н-с2-он 6сн2он
I
4
но-с3-н ^
н s
с о
14 у v. ОН Н s \ ► Полимеризация
Н-Сб-ОИ ОН С3—С 2
н он
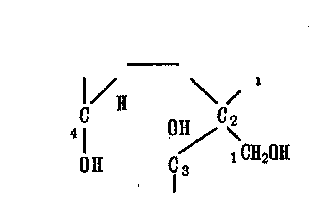
Глюкоза стереоспецифична, так что изменение взаимного располо-жения атомов Н и ОН приводит к образованию новых Сахаров с различными свойствами. Наряду с шестичленным циклом, образующимся при взаимодействии альдегидной группы Q с гидроксильной группой Сб, глюкоза может образовывать 5-членный цикл ~ фуранозу (фруктозу):
бСН20Н I
Н 5 С 0 qjw
н
фруктофураноза
Крахмал (полимер глюкозы в о-форме, в которой ОН-группы при Ci и Q>2 расположены по одну и ту же сторону от плоскости кольца)
303
запасается растениями в семенах и клубнях, а также в пшенице и кукурузе. Гликоген — крахмалоподобное вещество, синтезируемое в клетке. Он накапливается в мышцах и печени, поддерживает концентрацию глюкозы в крови. Молекулярная масса гликогена достигает 5 млн. Целлюлоза состоит из неразветвленных цепей, построенных из остатков глюкозы в /?-форме (ОН-1 и ОН-2 находятся по разные стороны плоскости пиранозы). Молекулярная масса полимеров целлюлозы достигает 500 000.
Белки представляют собой линейные полимеры аминокислот (амид-ная связь):
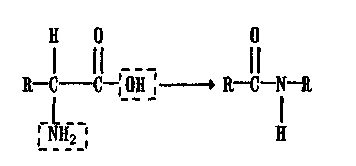
Белки гидролизуются с образованием аминокислот, которые затем дезаминируются в результате оксидазных реакций.
В конечном итоге глицерин, глюкозу и аминокислоты превращаются в пировиноградную кислоту, которая декарбоксилируется с преобразованием, как и при /3-окислении жирных кислот, в ацетил-КоА.
Ацетил-КоА служит исходным субстратом цикла трикарбоновых кислот (цикл Кребса); кроме того, ацетил-КоА служит для синтеза новых органических веществ.
Рис. 45. Схема внутриклеточного обмена веществ
Общая схема метаболизма органических веществ в клетке представлена на рис. 45.

304

Рис. 46. Схема трансформации карбоновых кислот в цикли Кребса
Цикл Кребса включает синтез из щавелевой кислоты и ацетил-КоА лимонной кислоты и последующее превращение лимонной кислоты до щавелевой с высвобождением восстановительных эквивалентов и СОг (рис. 46).
Из природных биополимеров особое значение имеют нуклеиновые кислоты, ДНК и РНК. Молекулярная масса ДНК лежит обычно в диапазоне 6—10 млн., но может достигать и 1010—1011. Если спираль ДНК растянуть в длину, она вытянется на несколько сантиметров. У РНК молекулярная масса меньше — 20—40 тыс.
Мономерное звено у этих полимеров — нуклеотид, содержащий остаток фосфорной кислоты, остаток сахара в фуранозной форме (5-членное кольцо) и остаток N-органического соединения с циклической структурой. Остаток сахара в РНК — рибоза, в ДНК — дезоксирибоза:
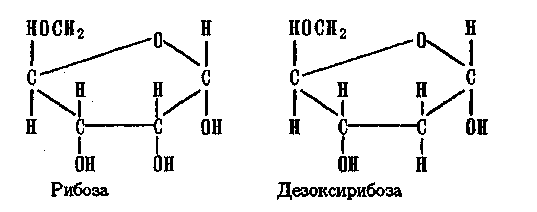
(В — остаток N-основания: аденин, гуанин, цитозин, тимин) 306
Полимеризация остатков рибозы происходит по типу конденсации аминокислот, но с образованием сложноэфирной фосфатной связи (молекула НгО отщепляется при комбинации ОН-группы сахара и ОН-группы фосфата):

Фактически образование биополимеров осуществляется за счет процессов конденсации с выделением НгО. С этим обстоятельством связана относительная легкость гидролиза большинства биополимеров под действием протолитических ферментов.