

Глава 3 круговорот веществ в биосфере
Устойчивость биосферы к воздействию внешних возмущающих факторов, к которым относится и человеческая деятельность, обусловлена существованием множества циклических процессов обмена химическими элементами между различными компонентами биосферы. Круговорот биогенных элементов в биосфере — углерода, кислорода, азота, серы и др. — представляет собой переход их из минеральной формы в живую материю и обратно за счет множества химических и биохимических превращений. Характерное- время биологического оборота запасов питательных веществ в окружающей среде оценивается в 10 лет.
§ 3.1. Круговорот кислорода, фотосинтез
В настоящее время в атмосфере содержится 1,2 «Ю15 т Ог- В результате фотосинтеза ежегодно образуется 2,3* 10й т сухого органического вещества и 2,5-1011 т Ог- Почти все это количество кислорода используется в процессах дыхания и ферментации органического вещества гетеротрофными, нефотосинтезирующими организмами, главным образом бактериями. При этом в атмосферу возвращается углекислый газ, использованный растениями при фотосинтезе.
Количество кислорода, не израсходованного гетеротрофными организмами, определяется массой органического вещества, поступающего в осадочные породы и выпадающего из глобального круговорота соединений углерода. Скорость накопления органического углерода в осадках оценивается величиной (1,4—3,0) * 107 т/год, что соответствует (5—8) • 107 т Ог, т.е. 0,4% фотосинтетического кислорода. Общие же запасы ископаемого углерода оцениваются в 1,25-Ю16 т (см. § 1.2).
Другой источник кислорода в атмосфере — процесс фотодиссоциации молекул воды — имеет пренебрежимо малое влияние на его общий баланс. Считается, что в ходе этой фото диссоциации образуется ~ 2-Ю6 т кислорода в год. Таким образом, образование и потребление кислорода происходит практически в замкнутом цикле фотосинтеза и микробиологической деструкции органического вещества в биосфере. Фактически состав атмосферы находится под контролем биоты — живого вещества планеты.
Биомасса живых организмов на Земле оценивается величиной 2-1012 т в расчете на сухой вес, причем более 90% ее приходится на долю фотосинтезирующих организмов, из которых 82% сосредоточено в лесах. Второй по величине резервуар живого вещества составляют почвенные микроорганизмы — бактерии, грибы, микроводоросли. Продукция их ненамного уступает продукции высших растений. Животное население планеты гораздо более разнообразно,, чем растительное. Общая же биомасса животных составляет около 2*109 т, т.е. всего около 0,1% биомассы высших растений. Соответственно вклад животных в биологический круговорот веществ пренебрежительно мал по сравнению с вкладом микроорганизмов и автотрофных растений.
В основе процессов жизнедеятельности лежит трансформация энергии солнечного излучения в энергию химических связей при фотосинтезе и в случае аэробных организмов — медленное "сжигание" органических веществ при окислении их кислородом воздуха в процессах дыхания. Цикл кислорода можно представить простой схемой:
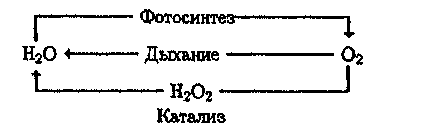
В биологическом круговороте кислорода, воды и углерода основная роль принадлежит фотохимическим процессам.
Особого успеха в осуществлении фотосинтеза достигли наземные растения. Не случайно для многих древних религий характерно поклонение дереву. Деревья играют планетарную роль в биохимическом круговороте воды. Вода, всасываемая корнями, поднимается по капиллярам ствола и ветвей дерева к листьям, в которых содержатся так называемые хлоропласты, где и протекает фотосинтез.
Хлоропласты — это специальные органеллы клетки. Типичная растительная клетка содержит 50—200 хлоропластов, каждый длиной около 1 мкм. Хлоропласты имеют систему внутренних мембран, так называемых ламелл, в которых локализованы фотосинтетические пигменты. Ламеллы образуют многослойные структуры, упакованные в пачки — граны.
Фотосинтезу в листьях сопутствует испарение воды с их поверхности (это явление называется транспирацией), осуществляемое через своеобразные "щели" (устьица, ширина которых меняется от 0 в зак-
85
рытом состоянии до 10 мкм в полностью раскрытом). На 1 мм2 поверхности листа может находиться несколько сотен таких устьиц — до 1% площади листа при их полном раскрытии. Вследствие транспирации листья не перегреваются на прямом солнечном свету. Например, береза за день испаряет в среднем 400 л воды.
Образование органического вещества из неорганической материи представляет собой первичную продукцию фотосинтеза. Ежегодно в процесс фотосинтеза на нашей планете вовлекается 38 км3 воды. Механизм фотосинтеза до конца еще не ясен.
Исследование фотосинтеза было фактически начато еще в 1650 г., когда естествоиспытатель Ван Гельмонт вырастил ивовое дерево из 5-фунтового саженца, посаженного в 200 фунтов почвы. Через 5 лет дерево весило 570 фунтов, а земля — 199 фунтов. Ван Гельмонт предположил, что увеличение веса дерева произошло за счет воды, которую он добавлял в почву.
Столетие спустя было обнаружено, что зеленые листья, помещенные в воду, под действием Солнца выделяют пузырьки газа, а Джозеф Пристли в 1771 г. доказал, что выделяемый зелеными растениями в дневное время газ не что иное, как 02.
Несколько позже было показано, что для выделения 02 растениям необходимы зеленые листья и свет. В 1782 г. Сенебье показал, что растениям для выделения Ог необходим углекислый газ, и это открытие привело к выводу, что С02 является источником всех органических веществ в растениях.
К середине прошлого столетия ученые пришли к выводу, что энергия, поступающая в виде солнечного света, запасается в форме химической энергии. И лишь в 20-е годы нашего столетия микробиолог Ван Ниль положил начало формированию современных представлений о фотохимических превращениях при фотосинтезе.
В природе обнаружены два основных типа фотосинтетических организмов.
Первый — фотосинтезирующие бактерии (пурпурные и зеленые), которые восстанавливают С02 до углеводородов, но не способны окислять воду до 02. В качестве доноров они используют не Н20, а органические и серные соединения (уксусная и пировиноградная кислоты, Н2) тиосульфата) и запасают относительно мало энергии. Многие фотосинтезирующие бактерии способны к фотоассимиляции N2.
Второй — зеленые растения и водоросли (красные, зеленые, сине-зеленые и др.), которые наряду с восстановлением С02 окисляют воду до 02. Ключевым процессом запасания солнечной энергии в виде химической является как раз окисление воды до 02.
Известны три молекулы (Н20, С02, N2), способные к образованию в 86
фотосинтетических процессах химических соединений, богатых свободной энергией. Ниже приведены фотосинтетические реакции с участием этих молекул с указанием количества запасаемой свободной энергии в расчете на перенесенный под действием света электрон:
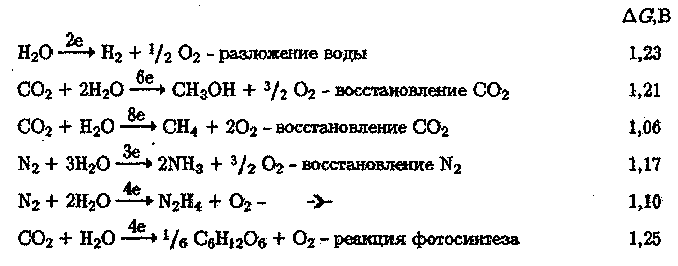
Окисление воды является наиболее фундаментальной реакцией. В каждой, фотосинтетической реакции вода участвует как восстановительный агент — донор Н.
Для образования одной молекулы Ог из двух молекул Н20 необходимо перенести четыре электрона, как и для восстановления СОг до уровня глюкозы:
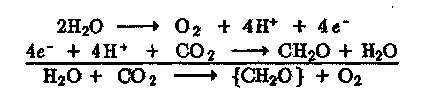
Природный процесс фотосинтеза протекает как бы в два этапа: первый -— образование Ог при поглощении четырех квантов света и восстановительных эквивалентов {2Н}:
![]()
второй — темновая стадия синтеза органического вещества:
![]()
Последовательность, в которой протекают фотохимические реакции процесса фотосинтеза, была изучена Хиллом и Вендалом в I960 г. Предложенная ими так называемая 2-схема согласуется со всеми известными на настоящее время данными (рис. 3).
87
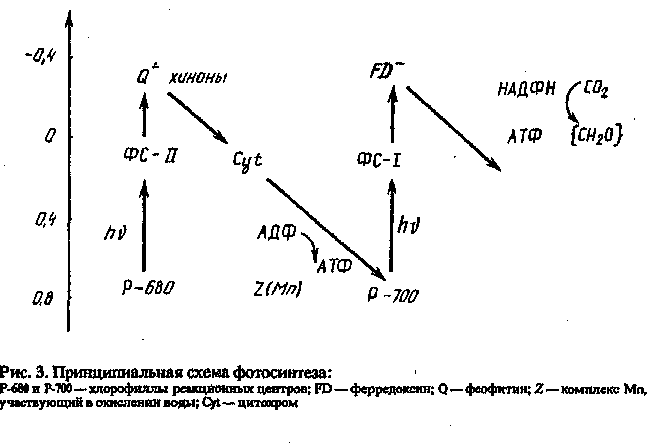
Эта схема состоит из двух основных частей: фотосистемы I (ФС-I) и фотосистемы II (ФС-П). ФС-I сопряжена с циклом Кальвина (рибуло-зодифосфатным циклом), в котором С02 восстанавливается до углеводов. Начинается этот цикл присоединением СОг к рибулозе-1,5-дифос-фату с образованием двух молекул 3-фосфоглицериновой кислоты:
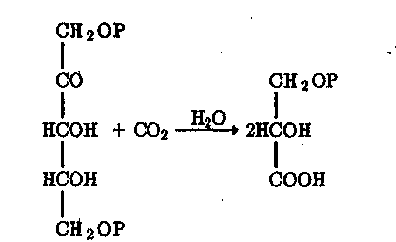
где Р — остаток фосфорной кислоты (—POj»").
Ё хлоропластах ФС-I и ФС-П. пространственно разделены. Каждая система содержит свой набор фотосинтетических пигментов, в которых основным компонентом является хлорофилл.
Молекулы хлорофилла представляют собой хромофоры, с помощью которых поглощается свет. В зеленых растениях содержится хлорофиллы an b, все различие которых заключается в замещении боковой группы ~СН3 (а) на группу —СНО (ft). Поглощение света в видимой
области спектра обусловлено наличием у молекул хлорофилла сильно сопряженной порфириновой системы. Хлорофилл поглощает свет в основном в синей (450 нм) и красной (650 нм) областях спектра, но отражает зеленый, желтый и оранжевый свет. Этим определяется характерный зеленый цвет растений.
После поглощения света возбужденная молекула хлорофилла может участвовать в процессах двух типов. Она может флуоресцировать или использовать энергию возбуждения для проведения каких-либо энергетически невыгодных химических реакций. Флуоресценция наблюдается лишь у хлорофилла а. Хлорофилл b в хлоропластах не флуоресцирует, он входит в состав светособирающей пигментной матрицы, помогающей улавливать и собирать свет. После фотовозбуждения хлорофилл b передает энергию возбуждения молекуле хлорофилла а — компоненту светособирающей матрицы с самой низкой энергией.
Кроме хлорофилла 6 в состав светособирающей матрицы входят каротиноидные пигменты и фикобилины. Функция фотосинтетических пигментов заключается в сборе квантов поглощаемого света и переносе энергии в реакционный центр, где происходит первичный перенос электрона, сопровождающийся разделением зарядов. Как правило, на каждый реакционный центр приходится 300 молекул пигмента. Средняя концентрация пигментов внутри светособирающей матрицы порядка ОД М.
Энергия, переносимая молекулами хлорофилла а, достигает реакционного центра, в котором электронно-возбужденная молекула (первичный донор электрона) переносит энергию к первичному акцептору Q с образованием продуктов электронного переноса.
Первичными донорами электронов в ФС-1 и ФОН в зеленых растениях служат пары (димеры) молекул хлорофилла в возбужденном состоянии (СЫ^).
Первичными акцепторами электронов служат мономерные молекулы хлорофилла а или феофитина (Q):
ОЫ*+ Q—КЩ + Q'
Образующиеся в реакционном центре ионы термодинамически нестабильны по отношению к обратному переносу электрона, но в результате вторичных процессов переноса электрона окислительные и восстановительные эквиваленты переносятся к центрам, находящимся на больших расстояниях. Вторичный перенос электрона представляет собой термический процесс, что является наиболее существенной особенностью природного фотосинтеза. Последовательность реакций фотосинтеза в ФС-И представлена на рис. 4.
№
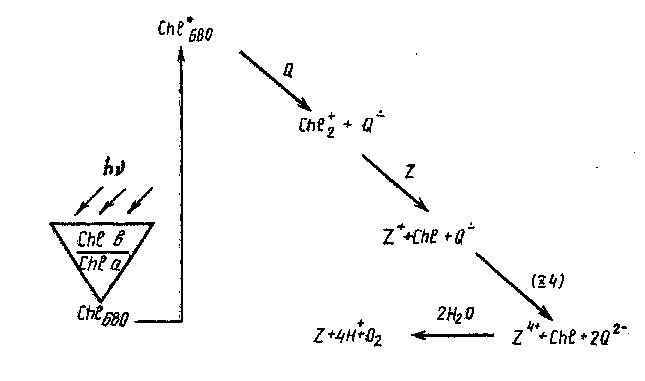
Рис. 4. Последовательность трансформации световой энергии в химическую в Ф-И
Катион-радикал димера хлорофилла окисляет воду в 4-электрон-ном процессе, включающем, по-видимому, марганец.
Если фотосинтезирующие организмы растут в отсутствие ионов Мп, то они теряют способность к выделению Оз- Встраивание Мп в дефектные клетки возвращает эту способность. х Максимальная активность достигается при включении одного иона Мп на 50—100 молекул хлорофилла а. В реакционном центре ФОН находится 4 атома Мп. Формальная схема фотокаталитического Электронного окисления воды может быть представлена в виде
4СЫ+ + Z—► 4СЫ2 + Z*+ Z*+ + 2Н20 —► Z + 4Н+ + 02
Используя импульсные лампы, с помощью чувствительных кислородных электродов было показано, что для выделения молекулы 02 требуется четыре кванта света.
Эффективность преобразования солнечной энергии в молекулярных системах природного фотосинтеза сравнительно высокая — 5—10%.
В биохимическом круговороте веществ синтезированные в процессе фотосинтеза органические вещества выполняют функцию строительного материала для обеспечения биологического каркаса всего "здания" жизни. В процессах жизнедеятельности часть органических веществ окисляется молекулярным кислородом с участием ферментов в качестве катализаторов: 90
{CH20} + 02 —► С02 + Н20
Химическая энергия, высвобождающаяся в этом процессе, расходуется на обеспечение энергией процессов синтеза органических соединений. Тем самым система фотосинтеза — окисление преобразует энергию солнечного излучения в химическую энергию процессов жизнедеятельности аэробных организмов.
Дыхание и фотосинтез — два противоположных процесса в природной среде, связанные с круговоротом кислорода и углерода. Углерод в биосфере занимает особое место, так как служит строительным материалом для органической жизни, источником энергии, высвобождаемой в процессе дыхания. Велика также роль неорганического углерода С02 в атмосфере — для регулирования климата и карбонатной системы, поддерживающей постоянство рН водной среды Океана.
Интенсивное сжигание ископаемого топлива, уничтожение лесов, разрушение почвенного покрова — все это приводит к участию человека в глобальных циклах круговорота углерода и кислорода (см. § 1.2).
Расходование полезных ископаемых — нефти, угля, газа — приводит к неизбежному истощению их запасов, горение — к продуктам окисления и в конечном итоге к образованию С02.
Хотя часть диоксида углерода вновь возвращается в органические соединения в процессе природного фотосинтеза зеленых растений или осаждается в виде океанических осадочных отложений, скорость индустриального расходования ископаемого топлива настолько велика, что накорление С02 в атмосфере не компенсируется его естественной убылью. За последние 150 лет количество сжигаемого топлива увеличивается ежегодно примерно на 4,3%.