

Эволюционное учение
.pdfважные видовые признаки. Для каждого вида свойственны свои границы пригодных для жизни условий, своя история возникновения (определяющая в значительной степени очертания и размер ареала), свои специфические взаимодействия с видами-конку- рентами и т. д. Поэтому любая характеристика вида будет неполной без указания на специфические особенности его ареала.
Однако существуют причины, по ко- |
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||
торым |
географический |
критерий |
не |
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||
может считаться достаточно универ- |
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||
сальным |
|
для |
характеристики |
видов |
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||
в природе. Во-первых, существует мно- |
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||
жество видов с практически совпа- |
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||
дающими |
|
ареалами. |
Например, |
ареал |
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||
кедрового |
|
|
стланника |
(Pinus |
pumila) |
|
|
|
|
|
|
|
|
|
|
|
|
||||||||
совпадает с ареалами березы Мид- |
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||
дендорфа |
|
|
(Betula middendorfii), дау- |
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||
рской лиственницы (Larix daurica) и |
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||
душистого |
тополя (Populus |
suaveolens). |
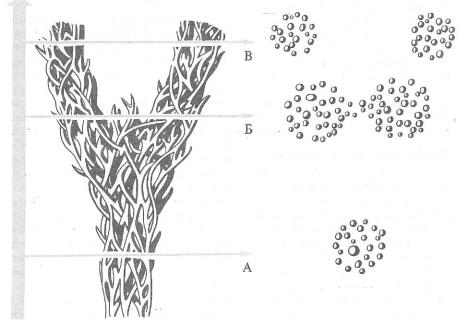
Рис. |
12.1. Схема интеграции генофондов от- |
|||||||||||||||||||||
Эти виды |
тесно |
связаны |
биогеоцено- |
||||||||||||||||||||||
дельных популяций в единый генофонд вида. |
|||||||||||||||||||||||||
тически, входят в состав одних и тех же |
|||||||||||||||||||||||||
Популяции — окружности |
различной |
|
величины, |
||||||||||||||||||||||
растительных сообществ, и их истори- |
|
||||||||||||||||||||||||
двойные линии — потоки генов между популяциями, |
|||||||||||||||||||||||||
ческое |
развитие |
шло, |
видимо, |
парал- |
|
|
|
пунктир—возникшая |
изоляция |
|
|
||||||||||||||
лельно, отражая общность историчес- |
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||
ких условий формирования определен- |
Возникает |
вопрос, |
а |
есть |
ли |
такой |
|||||||||||||||||||
ных природных |
зон. |
Во-вторых, |
для |
||||||||||||||||||||||
критерий? Есть, и он характеризует |
|||||||||||||||||||||||||
видов-космополитов, |
|
ареал |
которых |
||||||||||||||||||||||
|
вид и как всеобщую форму существо- |
||||||||||||||||||||||||
охватывает |
огромные |
пространства в |
|||||||||||||||||||||||
вания жизни на нашей планете, и |
|||||||||||||||||||||||||
биосфере |
|
Земли |
(в |
океане, |
например, |
||||||||||||||||||||
|
как |
качественный этап |
эволюционного |
||||||||||||||||||||||
дельфин |
|
косатка — Orcinus |
orca; |
на |
|||||||||||||||||||||
|
процесса. |
Основной |
критерий |
вида |
за- |
||||||||||||||||||||
суше — некоторые |
виды |
рода |
дрозо- |
ключается |
в |
его генетическом |
единстве. |
||||||||||||||||||
фил) , характеристика ареала как видо- |
Единство вида как динамической сис- |
||||||||||||||||||||||||
вого признака |
также |
теряет |
смысл. |
||||||||||||||||||||||
темы |
в |
эволюции |
основано |
на |
воз- |
||||||||||||||||||||
В-третьих, |
|
определенность |
ареала |
от- |
|||||||||||||||||||||
|
можности нивелировок различий, воз- |
||||||||||||||||||||||||
сутствует |
у |
некоторых |
быстрорасселя- |
||||||||||||||||||||||
никающих в популяциях путем скре- |
|||||||||||||||||||||||||
ющихся |
видов, |
поскольку |
очертания |
||||||||||||||||||||||
щивания. Как бы ни были изолированы |
|||||||||||||||||||||||||
ареала |
у |
них |
изменяются |
очень |
быст- |
||||||||||||||||||||
отдельные популяции и подвиды, поток |
|||||||||||||||||||||||||
ро и |
непредсказуемо |
(в |
последние |
||||||||||||||||||||||
генетической информации между ними |
|||||||||||||||||||||||||
столетия |
|
часто |
в |
связи |
с |
антропо- |
|||||||||||||||||||
|
всегда |
существует. |
Даже единичные |
||||||||||||||||||||||
генными изменениями ландшафтов или |
|||||||||||||||||||||||||
миграции отдельных особей из одной |
|||||||||||||||||||||||||
вселениями, |
как, |
например, |
для |
домо- |
|||||||||||||||||||||
популяции в другую, из зоны обита- |
|||||||||||||||||||||||||
вой мухи, |
|
городского |
воробья, многих |
||||||||||||||||||||||
|
ния |
|
одного |
подвида |
в |
зону |
обита- |
||||||||||||||||||
рудеральных растений |
и т. д.). |
|
|
|
|||||||||||||||||||||
|
|
ния другого, повторяясь на протяже- |
|||||||||||||||||||||||
Итак, |
|
и |
географический |
критерий |
нии |
|
сотен |
и |
тысяч |
поколений, |
созда- |
||||||||||||||
|
ют |
|
поток |
|
генов. |
Это |
обеспечивает |
||||||||||||||||||
не может |
|
служить |
каким-либо |
уни- |
|
|
|||||||||||||||||||
|
интеграцию |
относительно |
изолирован- |
||||||||||||||||||||||
версальным |
«видовым» |
признаком. |
|||||||||||||||||||||||
ных |
|
генофондов отдельных популяций |
|||||||||||||||||||||||
Генетическое |
единство — главный |
|
|||||||||||||||||||||||
(рис. |
12.1). |
|
|
|
|
|
|
|
|||||||||||||||||
критерий |
|
вида. |
Целостность |
вида. |
|
|
|
|
|
|
|
||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||
i 22 Часть 3. Учение о микроэволюции
Конечно, при отсутствии обмена гамет (у агамных, облигатнопартеногенетических и самооплодотворяющихся форм) не возникает такого генетического единства особей, что создает объективные трудности при разграничении видов.
Единство вида не нарушается даже и в том случае, если иногда по ка- ким-то причинам границы между соседними видами оказываются нечеткими. Сейчас в разных группах таких случаев вскрывается все больше. Это связано с тем, что особи близких видов в определенных условиях иногда могут успешно скрещиваться друг с другом и давать плодовитое потомство в некоторых комбинациях. В таких случаях оказывается, что, за исключением некой гибридной зоны, между такими видами нарушения строения обоих видов как самостоятельных генетических систем не происходит: виды оказываются не генетически закрытыми системами, как представлялось еще в середине XX в., а генетически устойчивыми системами.
Единство и эволюционная «судьба» вида определяют его специфическое место в биогеоценозах. В природе нет двух видов с одинаковым набором адаптаций: это разнообразие адаптаций делает каждый вид уникальным и определяет возможность формирования каждым видом своей экологической ниши} как суммы экологических ниш отдельных, составляющих вид популяций.
1 Не надо понимать экологическую нишу как некое пустое пространство, которое может быть занято тем или иным видом. Экологическая ниша — это как бы «профессия» данного вида, она возникает и исчезает одновременно с возникновением и исчезновением вида. Вне вида экологическая ниша не может существо-
вать. |
Вне |
вида существуют большие природ- |
ные |
зоны |
(зона лесов, пустынь, литораль |
и т. п.), формирующие соответствующие экологические ниши в результате развития тех или иных специальных адаптаций у определенных видов.
12.4. Использование понятия «вида» у агамных и облигатнопартеногенетических форм и в палеонтологии
Еще недавно формулировка понятия «вид» могла быть использована лишь по отношению к половым, перекрестнооплодотворяющимся формам (поскольку именно критерий скрещи-
ваемости |
являлся существеннейшим). |
|
К агамным, |
облигатнопартеногенети- |
|
ческим |
и |
самооплодотворяющимся |
формам, которые встречаются в природе в основном среди низших растений и микроорганизмов, это понятие трудно приложимо. Но виды существу-
ют и у таких форм. |
Сейчас, когда |
вид определяется не |
как генетичес- |
ки замкнутая, а как генетически устойчивая система, приложимость концепции вида к агамным и облигатнопартеногенетическим формам становится особенно ясной. У таких форм видом надо считать систему близких биотипов — группу фенотипически сходных особей, обладающих близкородственным генотипом, занимающих общий ареал и связанных общностью эволюционной судьбы. Эта общность определяется однообразием их генотипа. Образно говоря, агамные формы можно представить себе как одну огромную особь, состоящую из множества мелких однородных частей.
По-видимому, и агамные, и облигатнопартеногенетические формы формируют своеобразные генетические системы, устойчивость которых должна поддерживаться каким-то образом. Так, у бактерий нет механизмов генетической рекомбинации, связанных с половым процессом (мейозом, образованием и слиянием гамет). Но у прокариот выработались оригинальные способы обмена генетической информацией. Они связаны с заражением бактериальных клеток умеренными (т. е. не вызывающими гибель клеток) вирусами и их производными — плазмидами. В кольцевую молекулу ДНК плазмиды может включиться кусок

Глава 12. Вид — основной этап э в о л ю ц и о н н о г о процесса
«хозяйской» ДНК. Этот ген может быть перенесен от одной бактерии к другой. Бактерии обмениваются генами устойчивости к антибиотикам. Это явление получило название латерального переноса генетической информации.
|
Агамное |
размножение, |
партеноге- |
|
|
|
|
|||||||||||||
нез |
|
и |
самооплодотворение, |
видимо, |
|
|
|
|
||||||||||||
предствляют вторичное явление в эво- |
|
|
|
|
||||||||||||||||
люции специализированных в этом от- |
|
|
|
|
||||||||||||||||
ношении групп, и никогда не являются |
|
|
|
|
||||||||||||||||
абсолютно |
|
облигатными |
|
(даже |
для |
|
|
|
|
|||||||||||
цианей |
и |
ряда |
бактерий |
первичность |
|
|
|
|
||||||||||||
и |
облигатность |
|
агамии |
|
нельзя |
счи- |
|
|
|
|
||||||||||
тать доказанной). Подавляющее же |
|
|
|
|
||||||||||||||||
большинство |
|
организмов — двуполые, |
|
|
|
|
||||||||||||||
перекрестнооплодотворяющие |
формы. |
|
|
|
|
|||||||||||||||
|
Принципиальные |
затруднения |
воз- |
|
|
|
|
|||||||||||||
никают |
при |
|
использовании |
|
понятия |
|
|
|
|
|||||||||||
«вид» в палеонтологии. Здесь иссле- |
|
|
|
|
||||||||||||||||
дователь всегда имеет дело с рас- |
|
|
|
|
||||||||||||||||
пределением |
|
разных |
форм |
не |
только |
|
|
|
|
|||||||||||
в пространстве |
(как |
в |
|
неонтологии), |
|
|
|
|
||||||||||||
но и во времени. Это обстоятельство |
|
|
|
|
||||||||||||||||
лишает смысла критерий непосред- |
|
|
|
|
||||||||||||||||
ственного, |
«кровного» |
|
родства. |
Как |
|
|
|
|
||||||||||||
известно, в палеонтологии при раз- |
|
|
|
|
||||||||||||||||
граничении форм используются прак- |
|
|
|
|
||||||||||||||||
тически лишь морфологические и ре- |
Рис. 12.2. Схема строения вида как многоуров- |
|||||||||||||||||||
конструкционно-биологические (остат- |
невой системы (по А. С. Баранову, 1986) |
|||||||||||||||||||
ки жизнедеятельности, следы, капроли- |
|
|
|
|
||||||||||||||||
ты |
и |
т. |
п.) |
|
критерии. |
Разграничение |
го вида распадается на относительно |
|||||||||||||
форм видового ранга в палеонтологии |
изолированные группы особей. Внут- |
|||||||||||||||||||
оказывается возможным пока лишь в |
ри популяции можно всегда выделить |
|||||||||||||||||||
крайне редких последовательно захо- |
недолговечные группы особей, объе- |
|||||||||||||||||||
роненных на одном и том же месте |
диненных |
более тесным генетическим |
||||||||||||||||||
многочисленных по числу особей сери- |
родством. У животных такие группы |
|||||||||||||||||||
ях материала с заметными морфоло- |
часто называют демами, у растений — |
|||||||||||||||||||
гическими изменениями в разных гори- |
биотипами. |
|
|
|||||||||||||||||
зонтах. |
Поэтому |
понятие |
«вид» |
не- |
|
Если рассматривать внутривидовые |
||||||||||||||
применимо в палеонтологии. Для обоз- |
|
|||||||||||||||||||
структуры выше популяционного уров- |
||||||||||||||||||||
начения |
|
отрезка |
филогенетического |
|||||||||||||||||
|
ня, |
то у животных |
таковыми оказы- |
|||||||||||||||||
ствола, |
эквивалентного |
виду |
неонтоло- |
|||||||||||||||||
ваются расы, подвиды, а у растений — |
||||||||||||||||||||
гии, в палеонтологии |
сейчас |
все |
шире |
|||||||||||||||||
экотипы и подвиды. Единодушие в тер- |
||||||||||||||||||||
используется |
понятие |
фратрии |
(фи- |
|||||||||||||||||
минологии |
в этой области отсутствует |
|||||||||||||||||||
лу ма) |
или |
же |
пользуются |
не видо- |
||||||||||||||||
не |
только |
между |
ботаниками, зоо- |
|||||||||||||||||
выми, а лишь родовыми определениями. |
||||||||||||||||||||
логами, микологами и микробиологами, |
||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
12,5. |
Структура |
вида |
|
|
но и среди специалистов внутри |
||||||||||||
|
|
|
|
|
каждой из названных дисциплин. Од- |
|||||||||||||||
|
При описании элементарной эволю- |
нако при этом все без исключения |
||||||||||||||||||
ционной единицы — популяции (см. гл. |
биологи, связанные с изучением видов, |
|||||||||||||||||||
8) |
было показано, что население любо- |
согласны, что любой вид — это не конг- |
||||||||||||||||||
ломерат |
|
популяций, а |
сложная |
сис- |
видов |
и процесса |
видообразования |
|
тема |
с |
многоуровневой |
иерархией |
в природе. Для наблюдателя в природе |
||||
биохорологических групп |
(рис. |
12.2). |
любой |
вид как |
бы «рассыпается» |
|||
Сэволюционно-генетической точки на множество составляющих его попу-
зрения важно подчеркнуть, что эта |
ляций, каждая из которых входит в |
||||||||||||||||||||||||||
система интегрирована время от вре- |
свои конкретные биогеоценозы, зани- |
||||||||||||||||||||||||||
мени происходящим генетическим об- |
мает определенное место в цепях |
||||||||||||||||||||||||||
меном |
между |
группами. |
|
|
|
|
|
питания и потоках энергии. Но эти, |
|||||||||||||||||||
12.6. Вид — качественный этап |
|
казалось |
бы, |
дискретные, |
совершенно |
||||||||||||||||||||||
|
самостоятельные, с собственной эволю- |
||||||||||||||||||||||||||
|
эволюционного |
процесса |
|
|
ционной судьбой популяции невидимы- |
||||||||||||||||||||||
Несмотря на различное морфофи- |
ми,но теснейшими генетическими «ни- |
||||||||||||||||||||||||||
зиологическое «содержание» вида раз- |
тями» |
связаны |
друг |
с |
другом |
и |
|||||||||||||||||||||
ных |
групп |
организмов, |
вид — цент- |
время |
от |
времени |
(пусть |
даже |
не |
в |
|||||||||||||||||
ральный |
и |
главнейший, |
качественный |
каждом поколении, а через несколько |
|||||||||||||||||||||||
этап |
процесса |
эволюции. |
Вид |
может |
поколений) |
обмениваются |
генетичес- |
||||||||||||||||||||
и должен рассматриваться как ка- |
ким материалом в результате мигра- |
||||||||||||||||||||||||||
чественный этап эволюционного про- |
ции отдельных особей. Этот незначи- |
||||||||||||||||||||||||||
цесса, |
потому |
что |
это — наименьшая |
тельный на первый взгляд поток генов |
|||||||||||||||||||||||
неделимая генетически устойчивая сис- |
цементирует вид в единую сложную |
||||||||||||||||||||||||||
тема в живой природе. Основной био- |
иерархическую |
систему |
|
популяций, |
|
не |
|||||||||||||||||||||
логический смысл вида состоит в том, |
позволяет накопиться серьезным раз- |
||||||||||||||||||||||||||
что он образует защищенный генофонд |
личиям между популяциями и их груп- |
||||||||||||||||||||||||||
(благодаря |
развитию |
разнообразных |
пами. Если же эти связывающие |
||||||||||||||||||||||||
изолирующих |
механизмов, |
защищаю- |
отдельные популяции и группы попу- |
||||||||||||||||||||||||
щих |
его от |
возможного |
потока генов |
ляций эволюционно-генетические «мос- |
|||||||||||||||||||||||
из других генофондов). |
|
|
|
|
|
тики» рушатся и не восстанавли- |
|||||||||||||||||||||
Вид |
как качественный |
этап |
эво- |
ваются |
на |
протяжении |
длительного |
||||||||||||||||||||
периода |
(т. |
е. |
возникает |
непреодоли- |
|||||||||||||||||||||||
люционного процесса внутренне проти- |
|||||||||||||||||||||||||||
мый |
изоляционный |
барьер), |
то |
|
внутри |
||||||||||||||||||||||
воречив. С |
одной |
стороны, вид как ре- |
|
||||||||||||||||||||||||
одной |
генетически открытой |
популя- |
|||||||||||||||||||||||||
зультат эволюции целостен, приспособ- |
|||||||||||||||||||||||||||
ционной системы возникают две, |
каж- |
||||||||||||||||||||||||||
лен |
к данной |
среде, |
|
генетически |
обо- |
||||||||||||||||||||||
|
дая из которых оказывается генетичес- |
||||||||||||||||||||||||||
соблен |
от |
всех |
других |
таких |
же |
ви- |
|||||||||||||||||||||
ки устойчивой |
по |
отношению |
к |
дру- |
|||||||||||||||||||||||
дов, |
стабилен; |
с |
другой |
стороны, |
как |
||||||||||||||||||||||
гой. |
Так, |
с |
возникновением |
|
нового |
||||||||||||||||||||||
этап |
эволюционного процесса он дина- |
|
|||||||||||||||||||||||||
вида |
завершается |
один |
этап |
|
гранди- |
||||||||||||||||||||||
мичен, |
имеет |
расплывчатые |
границы, |
|
|||||||||||||||||||||||
озного |
и |
постоянно текущего |
в |
живой |
|||||||||||||||||||||||
лабилен. |
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||
|
|
|
|
|
|
|
|
|
|
природе процесса эволюции и начи- |
|||||||||||||||||
Эта |
противоречивая, |
диалектичес- |
|||||||||||||||||||||||||
нается новый этап. Поэтому и можно |
|||||||||||||||||||||||||||
кая, по существу, специфика вида — |
сказать, |
что |
любой |
вид — качествен- |
|||||||||||||||||||||||
объективная реальность, которую необ- |
ный |
|
этап |
|
эволюционного |
процесса. |
|||||||||||||||||||||
ходимо учитывать при любом изучении |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||
|
|
|
Итак, |
можно |
сделать |
вывод, |
что |
основной |
критерий |
|
|
|
|
||||||||||||||
|
|
вида — генетическое единство, существование вида как еди- |
|
|
|
|
|||||||||||||||||||||
|
|
ной интегрированной в процессе эволюции |
генетической |
|
|
|
|
||||||||||||||||||||
|
|
системы. |
Любой |
вид |
представляет |
собой |
репродуктив- |
|
|
|
|
||||||||||||||||
|
|
но |
изолированный |
|
генофонд. |
Эта |
|
интегрированная |
|
сис- |
|
|
|
|
|||||||||||||
|
|
тема |
всегда |
занимает |
в |
|
биосфере |
определенное, |
прису- |
|
|
|
|
||||||||||||||
|
|
щее |
только |
данному |
виду |
место. |
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||
Глава 1 3
Видообразование —• результат микроэволюции
Под |
|
влиянием |
давления |
различ- |
и давать |
плодовитое |
потомство |
(т. е. |
||||||
ных элементарных эволюционных фак- |
пока |
существует |
поток |
генетической |
||||||||||
торов внутри видового ареала то в од- |
информации между разными популя- |
|||||||||||||
ной, то в другой популяции возни- |
циями внутри вида), вид остается еди- |
|||||||||||||
кают устойчивые изменения генотипи- |
ным |
как |
сложная |
интегрированная |
||||||||||
ческого |
состава популяций. Некоторые |
система. Однако в результате возник- |
||||||||||||
из этих |
элементарных |
эволюционных |
новения |
сильного |
давления |
изоляции |
||||||||
явлений могут в дальнейшем углуб- |
этот |
поток |
генетической |
информации |
||||||||||
ляться. При более сильной изоляции |
может прерваться. Тогда оказавшиеся в |
|||||||||||||
такие |
эволюционные |
явления |
могут |
изоляции |
|
части |
видового |
населения, |
||||||
накапливаться в популяциях под дей- |
накопив изменения под влиянием дей- |
|||||||||||||
ствием |
отбора. |
|
|
|
ствующих |
эволюционных |
факторов, |
|||||||
|
|
|
|
|
|
могут |
перестать |
скрещиваться |
при |
|||||
13.1. |
Видообразование — источник |
последующих встречах, станут гене- |
||||||||||||
тически |
самостоятельными. |
|
|
|||||||||||
возникновения |
многообразия |
Возникновение |
такой |
|
изоляции |
|||||||||
|
|
в живой природе |
|
|
||||||||||
|
|
|
между разными частями видового на- |
|||||||||||
|
|
|
|
|
|
|||||||||
Пока особи из разных популяций |
селения |
означает |
разделение одного |
|||||||||||
внутри вида хоть изредка могут скре- |
вида |
на |
два — процесс видообразова- |
|||||||||||
щиваться |
в природе |
друг с |
другом |
ния. |
Схематично |
этот |
процесс |
изо- |
||||||
Рис. 13.1. Схема видообразования. Каждая отдельная веточка — популяция. А— уровень исходного единого вида; Б—момент незавершенного разделения видов; В—два новых вида (по Ф. Г. Добржанскому, 1957)
|
|
|
|
|
|
|
|
|
|
|
i 22 Часть 3. Учение о микроэволюции |
|
|
|
|
|
|
|
|
|
|
||||||||||
бражен на рис. 13.1. Итак, |
|
видооб- |
Америку — с |
другой |
(рис. |
|
13.2). Не- |
||||||||||||||||||||||||
разование |
— |
|
это разделение |
(во |
време- |
сколько сотен тысяч лет назад в рай- |
|||||||||||||||||||||||||
ни |
и |
пространстве) |
|
прежде |
|
единого |
оне |
современного |
Берингова |
|
пролива |
||||||||||||||||||||
вида на два или несколько. Другими |
обитала предковая форма этих чаек. |
||||||||||||||||||||||||||||||
словами, |
видообразование |
— |
это |
разде- |
Впоследствии |
они |
(биологически свя- |
||||||||||||||||||||||||
ление |
генетически |
открытой |
|
системы |
занные либо с побережьями морей, |
||||||||||||||||||||||||||
(какими являются по отношению друг |
либо с крупными внутриконтиненталь- |
||||||||||||||||||||||||||||||
к другу популяции и их |
группы |
ными |
водоемами) |
начали |
распростра- |
||||||||||||||||||||||||||
внутри вида), на генетически закрытые |
няться |
на |
восток |
|
и |
|
запад, |
|
образо- |
||||||||||||||||||||||
(или обязательно устойчивые) систе- |
вав к нашему времени две непре- |
||||||||||||||||||||||||||||||
мы. |
Видообразование |
происходит в |
рывные цепи подвидов. Особи всех со- |
||||||||||||||||||||||||||||
результате постоянно |
|
совершающихся |
седних подвидов скрещиваются в при- |
||||||||||||||||||||||||||||
внутри вида процессов микроэволюции. |
роде |
и |
дают |
|
плодовитое |
|
потомство. |
||||||||||||||||||||||||
Все |
хорошо |
изученные |
микроэво- |
В |
районе |
же |
Северного |
|
и |
|
Балтий- |
||||||||||||||||||||
ского морей произошла |
встреча конеч- |
||||||||||||||||||||||||||||||
люционные процессы протекают в сово- |
|||||||||||||||||||||||||||||||
ных звеньев восточной и западной цепи |
|||||||||||||||||||||||||||||||
купностях скрещивающихся |
и |
генети- |
|||||||||||||||||||||||||||||
подвидов. |
Накопленные |
в |
|
процессе |
|||||||||||||||||||||||||||
чески |
перемешивающихся |
|
особей. |
|
|||||||||||||||||||||||||||
|
микроэволюции |
отдельных |
|
подвидов |
|||||||||||||||||||||||||||
Только |
в такой |
системе |
возможно |
об- |
|
||||||||||||||||||||||||||
различия в биологии |
(особенности об- |
||||||||||||||||||||||||||||||
разование |
бесчисленного |
множества |
|||||||||||||||||||||||||||||
раза жизни, некоторые |
|
морфологичес- |
|||||||||||||||||||||||||||||
различных |
генетических |
|
комбинаций, |
|
|||||||||||||||||||||||||||
|
кие |
|
особенности |
и |
др.) |
|
оказались |
||||||||||||||||||||||||
являющихся |
основой для |
|
эффективно- |
|
|
||||||||||||||||||||||||||
|
достаточными |
для возникновения двух |
|||||||||||||||||||||||||||||
го действия |
естественного |
отбора. Од- |
|||||||||||||||||||||||||||||
новых |
видов. |
Если |
по |
каким-либо |
|||||||||||||||||||||||||||
нако |
скрещивания |
и |
происходящая |
||||||||||||||||||||||||||||
причинам |
непрерывная |
|
цепь |
подвидов, |
|||||||||||||||||||||||||||
вследствие |
этого |
нивелировка |
затруд- |
|
|||||||||||||||||||||||||||
связывающих |
сегодня |
|
эти |
|
не |
вполне |
|||||||||||||||||||||||||
няют эволюционную дифференцировку |
|
|
|||||||||||||||||||||||||||||
разделенные |
виды, |
|
где-нибудь |
разор- |
|||||||||||||||||||||||||||
более |
или |
|
менее |
крупных |
|
совокуп- |
|
||||||||||||||||||||||||
|
|
вется, |
то |
возникнут |
|
два |
|
самостоя- |
|||||||||||||||||||||||
ностей особей (популяций и их групп) |
|
|
|||||||||||||||||||||||||||||
тельных |
вида. |
Сейчас |
|
же |
видообра- |
||||||||||||||||||||||||||
в пределах |
одного вида. |
|
Образование |
|
|||||||||||||||||||||||||||
|
зование |
в |
этой |
группе |
|
чаек |
находится |
||||||||||||||||||||||||
нового |
вида |
создает |
|
обычно |
непрео- |
|
|||||||||||||||||||||||||
|
как |
|
бы |
|
«в |
процессе |
становления» |
||||||||||||||||||||||||
долимые |
в |
природных |
условиях |
изо- |
|
|
|||||||||||||||||||||||||
(in |
statu |
nascendi), |
давая нам возмож- |
||||||||||||||||||||||||||||
ляционные барьеры, которые позволя- |
|||||||||||||||||||||||||||||||
ность наблюдать эволюцию в действии. |
|||||||||||||||||||||||||||||||
ют сохранять специфические адапта- |
|||||||||||||||||||||||||||||||
ции |
каждого |
вида |
и |
в конечном |
ито- |
|
Видообразование |
в |
|
группе |
австра- |
||||||||||||||||||||
ге |
определяют |
возможность |
сохране- |
|
|
||||||||||||||||||||||||||
лийских мухоловок. В настоящее время |
|||||||||||||||||||||||||||||||
ния |
и |
увеличения |
многообразия |
про- |
|||||||||||||||||||||||||||
в Австралии распространены два близ- |
|||||||||||||||||||||||||||||||
явления |
живого |
на |
|
нашей |
планете. |
||||||||||||||||||||||||||
|
ких вида мухоловок рода Pachycephala |
||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||
13,2. |
Примеры видообразования |
(P. rufogularis и P. inornata). В ре- |
|||||||||||||||||||||||||||||
зультате тщательного анализа палео- |
|||||||||||||||||||||||||||||||
|
В настоящее время изучены раз- |
географии и палеоклиматологии уда- |
|||||||||||||||||||||||||||||
личные пути возникновения новых ви- |
лось восстановить возможный путь об- |
||||||||||||||||||||||||||||||
дов |
в |
процессе |
микроэволюции. |
|
|
разования такой странной йа первый |
|||||||||||||||||||||||||
|
Видообразование |
в |
цепи |
подвидов |
взгляд ситуации, когда один из близ- |
||||||||||||||||||||||||||
больших чаек. По побережьям Бал- |
ких видов обитает лишь на неболь- |
||||||||||||||||||||||||||||||
тийского |
и |
|
Северного |
морей |
живут |
шом пространстве, занимаемом другим |
|||||||||||||||||||||||||
не |
скрещиваясь |
два |
вида |
|
крупных |
видом (рис. 13.3). Вначале существо- |
|||||||||||||||||||||||||
чаек — серебристая |
чайка |
(Larus |
аг- |
вал лишь один предковый вид, имев- |
|||||||||||||||||||||||||||
gentatus) |
и |
клуша-хохотунья |
(L. |
fus- |
ший широкий ареал (рис. 13.3,/). В |
||||||||||||||||||||||||||
cus). Эти два самостоятельных вида |
связи с наступлением засушливого пе- |
||||||||||||||||||||||||||||||
объединяются друг с другом через |
риода, изменений в растительности и |
||||||||||||||||||||||||||||||
непрерывную цепь подвидов, охваты- |
смещении дождевых зон возникла и |
||||||||||||||||||||||||||||||
вающих Северную Евразию, с одной |
развилась значительная изоляция меж- |
||||||||||||||||||||||||||||||
стороны, |
и |
|
Гренландию |
|
и |
Северную |
ду |
западной |
и |
восточной |
частью ви- |
||||||||||||||||||||
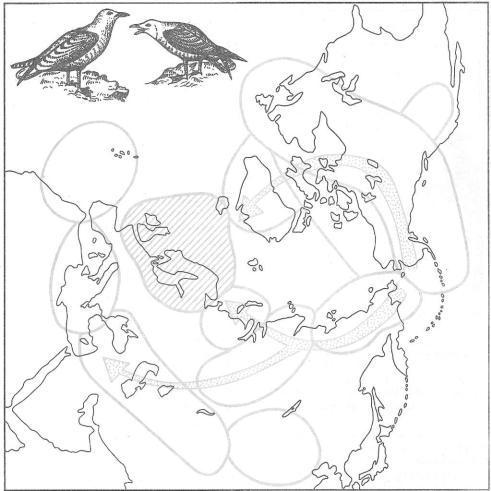
Рис. 13.2. Цепь подвидов больших |
чаек: серебристая—клуша-хохотунья |
(Larus argentatus— |
|||||||||||
L. fuscus) |
(по H. В. Тимофееву-Ресовскому и |
др., 1977). |
|
|
|
|
|
||||||
В ы д е л е н ы |
а р е а л ы отдельных |
подвидов. |
К р а й н и е |
з в е н ь я цепи |
в районе |
с е в е р о - з а п а д н о й |
Европы |
||||||
(двойная |
ш т р и х о в к а ) |
о б и т а ю т |
совместно |
и не |
с к р е щ и в а ю т с я в |
природе . |
Стрелки — основные на- |
||||||
п р а в л е н и я |
р а с с е л е н и я |
вида |
из |
вероятного Берингийского центра |
п р о и с х о ж д е н и я . |
В |
последнее в р е м я |
||||||
|
|
н е к о т о р ы м |
подвидам, в ы д е л е н н ы м |
на схеме, п р и д а е т с я видовой статус |
|
|
|||||||
дового |
населения (рис. 13.3, 2, 3). |
обретшим какие-то специфические осо- |
|||||||||||
Существуя в несколько иных усло- |
бенности. Различия между этими преж- |
||||||||||||
виях и при отсутствии нивелировок |
де генетически близкими формами ока- |
||||||||||||
достигаемых различий |
(при отсутствии |
зались настолько значительны, что оп- |
|||||||||||
скрещивания) |
две |
формы |
приобрели |
ределили возможность их самостоя- |
|||||||||
видовую |
самостоятельность |
(рис. |
тельного существования без скрещи- |
||||||||||
13.3,4). Затем вновь получила рас- |
вания. В сравнительно недавнее время |
||||||||||||
пространение сравнительно влаголюби- |
в результате нового иссушения клима- |
||||||||||||
вая растительность и западный вид |
та вновь возникла изоляция между |
||||||||||||
смог расселиться на восток, где он |
западной и восточной частями насе- |
||||||||||||
встретился с |
родственным |
видом, при- |
ления одного из |
видов |
(рис. |
13.3,5). |
|||||||
ммпммяяяретмяя
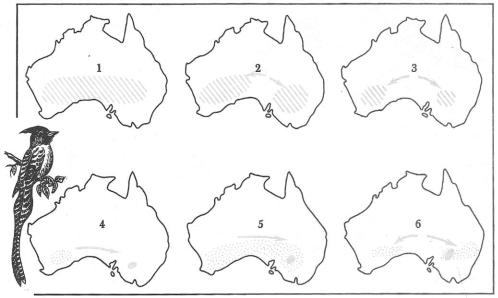
Рис. 13.3. Видообразование в группе австралийских мухоловок (Pachycephala) (из Э. Майра,
|
|
|
|
|
|
|
1968). |
|
|
|
|
|
|
||
Современное |
распространение показано |
на карте |
6. Расширения и |
сокращения ареалов |
связаны |
||||||||||
с изменениями в |
растительности, вызванными |
в свою очередь историческими смещениями |
|||||||||||||
|
|
|
дождевых зон в разные периоды. 1 — 6—этапы развития современных форм |
|
|
||||||||||
13.3. Основные пути и способы |
вида |
(дивергентное |
видообразование). |
||||||||||||
|
|
видообразования |
|
|
Наконец, новый вид может возник- |
||||||||||
|
|
|
|
|
|
|
|
нуть в результате гибридизации двух |
|||||||
|
Видообразование может быть рас- |
уже |
существующих |
видов — гибридо- |
|||||||||||
смотрено в территориальном и фило- |
генное видообразование. Кратко опи- |
||||||||||||||
генетическом аспектах. Новый вид мо- |
шем эти основные пути видообразова- |
||||||||||||||
жет |
возникнуть |
из |
одной |
или |
груп- |
ния. |
|
|
|
|
|
|
|||
пы смежных популяций, расположен- |
Аллопатрическое |
видообразование |
|||||||||||||
ных на периферии ареала исходного |
(называемое |
иногда |
географическим) |
||||||||||||
вида. Такое видообразование называет- |
иллюстрируется |
приведенными |
выше |
||||||||||||
ся |
аллопатрическим |
(от греч. alios — |
примерами |
возникновения |
вида у |
||||||||||
иной и patris — родина). В других слу- |
больших чаек и в группе австралий- |
||||||||||||||
чаях новый вид может возникнуть |
ских мухоловок. При аллопатрическом |
||||||||||||||
внутри ареала исходного вида, как бы |
видообразовании |
новые виды |
могут |
||||||||||||
внутри вида; этот путь видообразования |
возникнуть путем фрагментации, рас- |
||||||||||||||
называется |
симпатрическим |
(от |
греч. |
падения ареала широко распространен- |
|||||||||||
syn — вместе и |
patris — родина). Но- |
ного родительского вида. Примером та- |
|||||||||||||
вый вид может возникнуть посред- |
кого процесса может быть возникнове- |
||||||||||||||
ством постепенного |
изменения |
одного |
ние |
видов |
ландыша (см. гл. 6). |
||||||||||
и того |
же |
вида |
во |
времени, |
Другой способ аллопатрического видо- |
||||||||||
без какой-либо дивергенции исходных |
образования — видообразование |
при |
|||||||||||||
групп. Такое видообразование назы- |
расселении исходного вида, в процес- |
||||||||||||||
вается |
филетическим. |
|
|
се которого все более удаленные от |
|||||||||||
Новый вид может возникнуть пу- |
центра расселения периферийные попу- |
||||||||||||||
ляции и их |
группы, |
интенсивно |
пре- |
||||||||||||
тем |
разделения |
единого |
предкового |
||||||||||||
|
|
|
|
|
|
|
|||||||||
образуясь в новых условиях, становятся родоначальниками видов. Примеры, аналогичные видообразованию в группе больших чаек, известны и для других птиц, некоторых рептилий, амфибий, насекомых.
В основе аллопатрического видообразования лежат те или иные формы пространственной изоляции, и этот путь видообразования всегда сравнительно медленный, происходящий на протяжении сотен тысяч поколений. Именно за такие длительные промежутки времени в изолированных частях населения вида вырабатываются те биологические особенности, которые приводят к репродуктивной самостоятельности даже при нарушении первичной изолирующей преграды. Аллопатрическое видообразование всегда связано с историей формирования видового ареала.
|
Симпатрическое |
|
видообразование. |
|
|
|
|
|
|
|
|
|
|||||||||
При |
видообразовании |
симпатрическим |
|
|
|
|
|
|
|
|
|
||||||||||
путем новый вид возникает внутри |
|
|
|
|
|
|
|
|
|
||||||||||||
ареала |
исходного |
вида. |
|
|
|
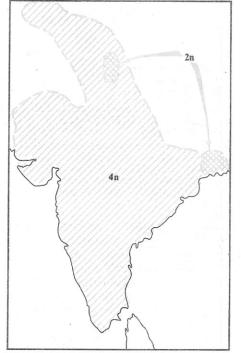
Рис. 13.4. Пример возникновения репродуктив- |
|||||||||||||||
|
Первый способ симпатрического ви- |
||||||||||||||||||||
|
ной изоляции при полиплоидизации в природе: |
||||||||||||||||||||
дообразования — возникновение |
но- |
обитающий по всему Индостану тетраплоидный |
|||||||||||||||||||
вых видов при быстром изменении |
вид растений Dicanthium annulatum несомненно |
||||||||||||||||||||
кариотипа, |
например |
при |
|
автополи- |
возник из диплоидной предковой формы, за- |
||||||||||||||||
плоидии. |
Известны |
группы |
близких |
нимающей сейчас небольшой и дизъюнктный |
|||||||||||||||||
|
ареал (по Н. Россу, |
1962) |
|
||||||||||||||||||
видов (обычно растений) с кратными |
|
|
|
|
|
|
|
|
|
||||||||||||
числами хромосом (см. рис. 6.28). |
|
|
|
|
|
|
|
|
|
||||||||||||
Так, например, в роде хризантем |
ловиях. Возникшие полиплоидные осо- |
||||||||||||||||||||
(Chrysanthemum) |
|
все |
|
формы имеют |
би могут давать жизнеспособное ло- |
||||||||||||||||
число |
хромосом, |
кратное |
9, |
18, |
27, |
томство лишь при скрещивании с осо- |
|||||||||||||||
36, 45..., 90. В родах табака |
(Ni- |
бями, несущими то же число хромосом |
|||||||||||||||||||
cotiana) и картофеля (Solanum) основ- |
(или при самоопылении). В течение |
||||||||||||||||||||
ное, исходное, число хромосом равно |
немногих |
поколений |
в |
том |
случае, |
||||||||||||||||
12, но имеются формы с 24, 48, 72 |
если |
полиплоидные |
формы |
|
успешно |
||||||||||||||||
хромосомами. В таких случаях можно |
проходят «контроль» естественного от- |
||||||||||||||||||||
предположить, |
что |
|
видообразование |
бора и оказываются лучше исходной |
|||||||||||||||||
шло путем автополиплоидии — посред- |
диплоидной, они могут распространить- |
||||||||||||||||||||
ством удвоения, утроения, учетвере- |
ся и сосуществовать совместно с поро- |
||||||||||||||||||||
ния и т. д. основного набора хромо- |
дившим |
их |
видом |
|
(рис. |
13.4) или, |
|||||||||||||||
сом предковых видов. Процессы поли- |
что |
бывает |
чаще, |
просто |
вытеснить |
||||||||||||||||
плоидизации |
хорошо |
|
воспроизводятся |
его. |
|
|
|
|
|
|
|
|
|||||||||
в |
эксперименте посредством |
задержки |
Полиплоидные формы, как правило, |
||||||||||||||||||
расхождения |
|
хромосом |
в |
мейозе, |
|||||||||||||||||
|
крупнее |
и |
способны |
существовать в |
|||||||||||||||||
в |
результате |
воздействия, |
например, |
||||||||||||||||||
более суровых физико-географических |
|||||||||||||||||||||
колхицином. Известно, что полиплоиды |
|||||||||||||||||||||
условиях. Именно |
поэтому |
в |
высоко- |
||||||||||||||||||
могут |
возникать |
и |
в |
природных |
ус- |
||||||||||||||||
горьях и |
в |
Арктике |
число |
|
полипло- |
||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||
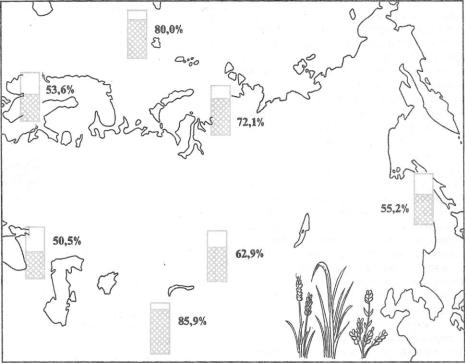
Часть 3. Учение о м и к р о э в о л ю ц и и
Рис. 13.5. Распространение полиплоидных видов цветковых растений (в процентах к общему числу видов) во флорах разных районов Евразии (по данным разных авторов из Н. В. Тимофеева-Ресовского и др., 1977)
идных видов растений резко увеличено |
которого |
|
полностью |
репродуктивно |
||||||||||
(рис. 13.5). Среди животных полипло- |
изолированы друг от друга, и, если |
|||||||||||||
идия при видообразовании играет не- |
условие отбора сохраняется, лишь во- |
|||||||||||||
сравненно меньшую роль, чем у рас- |
прос времени — когда эти формы при- |
|||||||||||||
тений, и |
во |
всех |
случаях |
связана |
обретут ранг новых видов. Аналогично |
|||||||||
с партеногенетическим способом раз- |
положение с яровыми и озимыми |
|||||||||||||
множения |
(например, |
у |
иглокожих, |
расами |
проходных |
рыб; возможно, |
||||||||
членистоногих, аннелид и других бес- |
что эти формы уже являются разными |
|||||||||||||
позвоночных) . |
|
|
|
|
|
|
видами, очень схожими морфологичес- |
|||||||
Второй способ симпатрического ви- |
ки, но |
изолированными |
генетически |
|||||||||||
дообразования |
— |
путем |
гибридизации |
(виды-двойники). |
|
|
|
|
||||||
с последующим удвоением числа хро- |
Особенность |
симпатрического пути |
||||||||||||
мосом — аллополиплоидия (см. ниже). |
видообразования — возникновение |
но- |
||||||||||||
Наконец, |
последним, |
достаточно |
вых видов, морфофизиологически близ- |
|||||||||||
изученным |
способом |
симпатрического |
ких к исходному виду. Так, при поли- |
|||||||||||
видообразования |
является |
возникно- |
плоидии |
увеличиваются |
размеры, |
но |
||||||||
вение новых форм в результате сезон- |
общий облик |
растений |
сохраняется, |
|||||||||||
ной изоляции. |
Известно |
существование |
как правило, неизменным; при хро- |
|||||||||||
ярко выраженных сезонных рас у |
мосомных |
перестройках |
наблюдается |
|||||||||||
растений, например у погремка Alec- |
та же картина; при экологической |
|||||||||||||
torolophus |
major |
(см. |
гл. |
10), ран- |
(сезонной) |
изоляции |
возникающие |
|||||||
нецветущие и |
позднецветущие |
формы |
формы |
также |
обычно |
оказываются |
||||||||