

Эволюционное учение
.pdf
|
|
|
Глава 9. Элементарные |
факторы |
эволюции |
|
131 |
||
химические |
вещества |
способны чаще |
признано явление колебания ее числен- |
||||||
вызывать определенные мутации. Нап- |
ности — волн жизни |
(популяционные |
|||||||
ример, окись этилена вызывает у ячме- |
волны, см. рис. 7.1). Популяционные |
||||||||
ня появление мутаций увеличенного ко- |
волны как общее свойство популяции |
||||||||
личества хлорофилла чаще, чем без- |
имеют важное общеэволюционное зна- |
||||||||
хлорофилловых |
мутаций, а небуларин |
чение |
как самостоятельный |
фактор |
|||||
дает противоположный эффект. Специ- |
эволюции. |
|
|
||||||
фичность действия мутагенов позволи- |
Периодические или |
апериодические |
|||||||
ла надеяться на возможность полу- |
колебания численности характерны для |
||||||||
чения |
направленных мутаций. |
Однако |
всех без исключения видов живых орга- |
||||||
даже |
действие |
самых |
специфических |
низмов. |
Конкретные |
причины |
таких |
||
мутагенов всегда дает спектр разнооб- |
флуктуаций могут быть бесконечно раз- |
||||||||
разных мутаций. Решать проблему нап- |
нообразны и откоситься к действию |
||||||||
равленного изменения |
наследственнос- |
как биотических, так и абиотических |
|||||||
ти приходится совершенно иным пу- |
факторов среды. Действие волн жизни |
||||||||
тем — с помощью «генетической инже- |
как эволюционного фактора предпола- |
||||||||
нерии»: посредством синтеза |
нужного |
гает неизбирательное, случайное унич- |
|||||||
гена и включения его в геном особи. |
тожение особей, благодаря чему ред- |
||||||||
Значение |
мутационного |
процесса |
кий перед колебанием численности ге- |
||||||
как эволюционного фактора. Эволю- |
нотип (аллель) может сделаться обыч- |
||||||||
ционное значение мутационного про- |
ным и подхвачен в дальнейшем естест- |
||||||||
цесса определяется прежде всего тем, |
венным |
отбором. |
|
|
|||||
что он постоянно поддерживает высо- |
Классификация |
популяционных |
|||||||
кую степень гетерогенности природ- |
волн. В природе встречается множество |
||||||||
ных популяций — основу для действия |
популяционных волн. Рассмотрим ос- |
||||||||
других факторов эволюции и прежде |
новные |
примеры. |
|
|
|||||
всего |
естественного |
отбора. |
Можно |
1. Периодические |
колебания |
чис- |
|||
сказать, |
что |
мутационный |
процесс |
— |
ленности |
короткоживущих |
организ- |
||||
это фактор-поставщик элементарного |
мов — характерны |
|
для |
большинства |
|||||||
эволюционного |
материала. |
|
|
насекомых, однолетних растений, боль- |
|||||||
Мутационный процесс ведет к воз- |
шинства |
грибов |
и |
микроорганизмов. |
|||||||
никновению части того «резерва» нас- |
В наиболее простом для анализа виде |
||||||||||
ледственной |
изменчивости, |
который |
эти сезонные колебания |
численности |
|||||||
определит |
в |
будущем возможность |
выражены у микроорганизмов: доста- |
||||||||
приспособления популяций к тем или |
точно вспомнить весенние и осенние |
||||||||||
иным изменениям условий среды. Об- |
волны простудных заболеваний, опре- |
||||||||||
разно говоря, груз мутации, непрерывно |
деляемые массовым размножением ря- |
||||||||||
пополняемый |
мутационным |
процес- |
да вирусов. У растений и животных |
||||||||
сом,— это цена, которую «платит» по- |
сезонные колебания численности не- |
||||||||||
пуляция сегодня за возможность сох- |
одинаково отражаются на разных воз- |
||||||||||
ранения в измененных условиях завтра, |
расти о-пол о в ы х |
группах |
популяции. |
||||||||
приобретения |
новых |
признаков |
и |
2. Непериодические |
колебания чис- |
||||||
свойств и освоения ранее недоступных |
ленности, зависящие от сложного со- |
||||||||||
условий существования. |
|
|
четания разных факторов. В первую |
||||||||
|
|
|
|
|
|
очередь они зависят от благоприятных |
|||||
9.3. Популяционные волны |
|
для данного вида |
(популяции) |
отноше- |
|||||||
|
ний в пищевых цепях: ослабление прес- |
||||||||||
как элементарный |
эволюционный |
са хищников для популяций жертв или, |
|||||||||
|
|
фактор |
|
|
например, увеличение кормовых ресур- |
||||||
При характеристике популяции как |
сов для популяций хищников. Обычно |
||||||||||
элементарной |
эволюционной |
единицы |
такие колебания численности касаются |
||||||||
одной из |
важных и |
постоянных черт |
не одного-двух, а многих видов живот- |
||||||||
5* |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
i 22 Часть 3. Учение о микроэволюции |
|
|
|
|
|
|
|
||||||||||
ных и растений в биогеоценозах и |
бирского шелкопряда в западно-сибир- |
||||||||||||||||||||||||||
порой ведут к коренным перестройкам |
ских |
лесных биоценозах. |
|
|
|
||||||||||||||||||||||
всего |
биогеоценоза. |
|
|
|
|
|
|
|
Эволюционное |
|
значение |
популя- |
|||||||||||||||
3. |
Вспышки |
|
численности |
видов |
в |
ционных волн. Если численность какой- |
|||||||||||||||||||||
новых районах, где отсутствуют их |
либо популяции резко идет на убыль, |
||||||||||||||||||||||||||
естественные враги. Примерами коле- |
то от многочисленной популяции слу- |
||||||||||||||||||||||||||
баний численности в XIX—XX вв. слу- |
чайно могут остаться немногие особи. |
||||||||||||||||||||||||||
жат кролики в Австралии, домовые во- |
Например, лесной пожар не затронул |
||||||||||||||||||||||||||
робьи в Северной Америке, канадская |
небольшой участок леса, где и сохрани- |
||||||||||||||||||||||||||
элодея, американская норка и ондатра |
лись остатки популяции короедов |
(био- |
|||||||||||||||||||||||||
в Евразии и др. В XVI—XVII вв. |
логические свойства оставшихся в жи- |
||||||||||||||||||||||||||
вспышки численности испытывали рас- |
вых короедов в данном случае не имели |
||||||||||||||||||||||||||
селявшиеся с мореплавателями по все- |
значения, |
|
их |
спасла |
случайность). |
||||||||||||||||||||||
му |
миру |
крысы |
(Rattus |
norvegicus). |
В популяции, пережившей катастрофи- |
||||||||||||||||||||||
Неимоверно |
увеличилась |
численность |
ческое сокращение численности, час- |
||||||||||||||||||||||||
домовой |
мухи |
|
(Musca |
domestica) |
в |
тоты |
генов |
(точнее, |
частоты |
аллелей) |
|||||||||||||||||
связи с распространением помоек и |
будут иными, чем во всей исходной |
||||||||||||||||||||||||||
гниющих |
|
пищевых |
запасов |
|
около |
популяции до начала спада численнос- |
|||||||||||||||||||||
поселений |
человека. |
|
|
|
|
|
|
ти. Если вслед за сокращением числен- |
|||||||||||||||||||
4. |
Резкие |
|
непериодические |
колеба- |
ности |
следует |
новая волна |
|
жизни и |
||||||||||||||||||
|
резкий |
подъем |
численности, |
|
то |
новой |
|||||||||||||||||||||
ния |
численности, |
связанные |
с |
природ- |
|
||||||||||||||||||||||
вспышке |
|
численности |
дает |
начало |
|||||||||||||||||||||||
ными |
«катастрофами» |
|
(разрушением |
|
|||||||||||||||||||||||
|
оставшаяся немногочисленная группа. |
||||||||||||||||||||||||||
биогеоценозов |
или |
целых |
ландшаф- |
||||||||||||||||||||||||
Генотипический |
состав |
этой |
группы и |
||||||||||||||||||||||||
тов) . Несколько засушливых лет могут |
|||||||||||||||||||||||||||
определит |
новую |
генетическую струк- |
|||||||||||||||||||||||||
быть причиной серьезных изменений в |
|||||||||||||||||||||||||||
туру всей популяции в период следую- |
|||||||||||||||||||||||||||
облике больших |
территорий |
(наступ- |
|||||||||||||||||||||||||
щего |
|
расцвета |
ее |
численности. |
При |
||||||||||||||||||||||
ление луговой растительности на боло- |
|
||||||||||||||||||||||||||
этом |
некоторые |
ранее |
присутствовав- |
||||||||||||||||||||||||
тистые |
места, |
|
увеличение |
площади |
|||||||||||||||||||||||
|
шие |
в |
малых концентрациях |
мутации |
|||||||||||||||||||||||
сухих лугов, выгорание большого числа |
|||||||||||||||||||||||||||
могут |
совершенно |
исчезнуть |
из |
попу- |
|||||||||||||||||||||||
торфяниковых |
залежей, |
сохранявших- |
|||||||||||||||||||||||||
ляции, |
а |
концентрация |
других |
мута- |
|||||||||||||||||||||||
ся |
на |
протяжении |
нескольких |
|
тысяч |
||||||||||||||||||||||
|
ций |
может |
случайно резко |
повысить- |
|||||||||||||||||||||||
лет |
как |
устойчивый |
биотип, |
и |
т. д.). |
||||||||||||||||||||||
ся (рис. |
|
9.3). |
В |
целом |
произойдет |
||||||||||||||||||||||
Виды |
с |
подвижными |
особями |
(круп- |
|
||||||||||||||||||||||
случайное |
|
колебание |
концентраций |
||||||||||||||||||||||||
ные |
млекопитающие, |
насекомые, |
пти- |
|
|||||||||||||||||||||||
разных |
генотипов |
и |
мутаций |
в |
попу- |
||||||||||||||||||||||
цы) |
или |
живущими |
в |
глубоких |
|
слоях |
|||||||||||||||||||||
|
ляции. |
В |
|
этом |
и |
состоит |
|
основное |
|||||||||||||||||||
почвы |
страдают |
меньше |
неподвижных |
|
|
||||||||||||||||||||||
эволюционное значение |
волн |
числен- |
|||||||||||||||||||||||||
и малоподвижных |
форм, живущих в |
||||||||||||||||||||||||||
ности. |
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||
лесу на почве. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||
Такие формы гибнут в огромном |
При |
определенных |
условиях это |
||||||||||||||||||||||||
числе |
(растения, |
моллюски, |
рептилии, |
случайное и кратковременное колеба- |
|||||||||||||||||||||||
амфибии и др.). На пожарищах хорошо |
ние численности может превратиться в |
||||||||||||||||||||||||||
известны вспышки численности иван- |
элементарное эволюционное явление — |
||||||||||||||||||||||||||
чая (Epilobium angustifolium) и свя- |
изменение на протяжении ряда поко- |
||||||||||||||||||||||||||
занного |
с |
|
ним |
сообщества |
|
насе- |
лений генотипического состава попу- |
||||||||||||||||||||
комых. |
|
|
|
|
|
|
|
|
|
|
|
|
ляции. |
|
|
|
|
|
|
|
|
|
|
||||
Масштабы |
|
колебаний |
численности |
Механизм действия популяционных |
|||||||||||||||||||||||
могут быть различными. Близкие, по- |
волн в эволюции иллюстрируется мо- |
||||||||||||||||||||||||||
видимому, |
к |
максимальным |
величины |
дельным примером. В закрытый ящик |
|||||||||||||||||||||||
колебаний численности в 1 млн. раз |
положено 500 белых и 500 черных |
||||||||||||||||||||||||||
,отмечены Ю. И. Новоженовым в |
шаров, что соответствует частоте генов |
||||||||||||||||||||||||||
Зауралье |
для |
майских |
жуков, |
и |
в |
в популяции: Р —0,50. Удалим из ящи- |
|||||||||||||||||||||
12 |
млн. |
раз — А. С. Исаевым для |
си- |
ка наугад 10 шаров |
(что соответствует |
||||||||||||||||||||||
|
|
abcdEf |
|
abcdef |
|
|
abcdef |
aBcdef |
( |
abcdef |
|
abcdef |
|
abcdef |
|
abcdef |
|
||||||||
|
|
abcdef |
|
abcdef |
|
|
abcdef |
abcdef |
|
abcdef |
|
abcdef |
|
Abcdef |
|
abcdef |
|
||||||||
|
|
abcdef |
|
abCdef |
|
|
abcdef |
abcdef |
|
abcdef |
|
aBcdef |
|
abcdef |
|
abcdef |
|
||||||||
|
|
abcdef |
|
abcdef |
|
|
abcdef |
abcdef |
|
abcdef |
|
abcdef |
|
abcdef |
|
abcdef |
|
||||||||
|
|
aBcdef |
|
abcdef |
|
|
abcdef |
abcdef |
|
abcdef |
|
abcdef |
|
abcdef |
|
abcDef |
|
||||||||
|
|
abcdef |
|
abcdef |
|
|
abcdef |
abcdef |
|
abcdef |
|
abcdef |
|
abcdef |
|
abcdef |
|
||||||||
|
|
abcdef |
|
abcdef |
|
|
aBcdef |
abcdef |
|
abcdef |
|
abcdef |
|
abcdef |
|
abcdef |
|
||||||||
|
|
abcdef |
|
abcdef |
|
|
abcdEf |
abcdef |
Г "abcdeF ~ ~ abcdef" ~j |
abcdef |
|
abcdef |
|
||||||||||||
|
|
abcdef |
|
abcdef |
|
|
abcdef |
abcdef |
|
abcdef |
|
abcdef |
J |
abcdef |
|
abcdef |
|
||||||||
|
|
abcdef |
|
abcdef |
|
|
abcdef |
abcdef |
|
abcdef |
|
abcdef |
|
abcdEf |
|
abcdef |
|
||||||||
|
|
abcdef |
|
abcdef |
|
|
abcdef |
abcdef |
|
abcdef |
|
abcdef |
|
abcdef |
|
abcdef |
|
||||||||
|
|
abcdef |
|
abcdef |
|
|
abcdef |
abcdef |
|
abcdef |
|
abcdef |
|
abcdef |
|
Abcdef |
|
||||||||
|
|
abcdef |
|
abcdef |
|
|
abcdef |
abcdef |
|
abcdef |
|
abcdef |
|
abcdef |
|
abcdef |
|
||||||||
|
|
abcdef |
|
abcdef |
|
|
abcdef |
aBcdef |
|
abcdef |
|
abcdef |
|
abcdef |
|
abcdef |
|
||||||||
|
|
aBcdef |
|
abcdef |
|
|
abcdef |
abcdef |
|
abcdEf |
abcdef |
|
abcdef |
|
abcdef |
|
|||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
—• |
|
|
|
Рис. 9.3. Схема действия популяционных волн: а, Ь, с, d, е, /—различные гены, в совокупности |
||||||||||||||||||||||||
|
обозначающие индивидуум, А, В, С, D, |
Е, |
F—соответствующие |
мутации, |
встречающиеся |
||||||||||||||||||||
|
в разной концентрации. Если в результате резкого изменения численности в популяции |
||||||||||||||||||||||||
|
сохранятся |
только выделенные пунктиром особи (в |
данном примере |
|
составляющие |
10% |
|
||||||||||||||||||
|
всего |
числа особей), |
то |
концентрация мутаций |
в популяции резко изменится (из Э. Бауэра, |
||||||||||||||||||||
|
|
|
|
|
|
|
|
|
Н. В. Тимофеева-Ресовского, 1943) |
|
|
|
|
|
|
|
|
|
|
||||||
«выжившим» |
5 |
диплоидным |
особям), |
особей не более 500. Именно в этих |
|||||||||||||||||||||
из |
них |
4 |
будут |
черными, 6— белыми |
условиях |
популяционные |
|
волны |
могут |
||||||||||||||||
|
(Р — 0,40 |
и |
Р—0,60). В следующий |
|
как бы подставлять под действие ес- |
||||||||||||||||||||
момент жизни нашей «популяции» об- |
тественного |
отбора |
редкие |
мутации |
|||||||||||||||||||||
щую численность |
«особей» |
увеличим |
(внося их в увеличенных концентра- |
||||||||||||||||||||||
в 100 |
раз |
(400 |
черных, 600 |
белых). |
циях |
в популяционный |
генофонд) или |
||||||||||||||||||
Если мы опять наугад удалим 10 шаров, |
устранять уже довольно обычные ва- |
||||||||||||||||||||||||
то велика вероятность, что у нас в |
рианты. |
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||
руках окажутся шары в соотношении, |
Итак, |
действие |
|
популяционных |
|||||||||||||||||||||
очень |
далеком |
от |
первоначального |
волн, так же как и действие мутацион- |
|||||||||||||||||||||
|
(например, 2 черных и 8 белых и соот- |
ного процесса, статистично и ненап- |
|||||||||||||||||||||||
ветственно Р —0,20 и Р—0,80). Это |
равленно |
(хотя природа действия фак- |
|||||||||||||||||||||||
соотношение частот в третьей наугад |
торов совершенно различна). Популя- |
||||||||||||||||||||||||
взятой выборке из средней величины |
ционные |
волны |
служат |
«поставщиком» |
|||||||||||||||||||||
«популяции» |
(200 черных и 800 белых) |
|
эволюционного |
|
материала. |
Действие |
|||||||||||||||||||
легко может случайно сдвинуться в сто- |
этого фактора трудно вычленить в |
||||||||||||||||||||||||
рону теперь уже преобладающих белых, |
«чистом |
виде»— очень |
|
мало |
точных |
||||||||||||||||||||
и «популяция» будет представлена поч- |
данных по оценке величины его давле- |
||||||||||||||||||||||||
ти лишь белыми шарами: велика ве- |
ния в природных популяциях. Теорети- |
||||||||||||||||||||||||
роятность того, что мы вытащим 9 бе- |
чески |
давление |
популяционных |
волн, |
|||||||||||||||||||||
лых и 1 черный (Р —0,10 и Р —0,90). |
особенно в малых по численности |
||||||||||||||||||||||||
Точно так же и в природных популя- |
популяциях, должно заметно превы- |
||||||||||||||||||||||||
циях случайные колебания частот ге- |
шать давление мутационного процесса. |
||||||||||||||||||||||||
нов |
могут |
быть |
причиной устранения |
Популяционные волны и мутацион- |
|||||||||||||||||||||
или |
резкого |
повышения концентрации |
|||||||||||||||||||||||
ный |
процесс |
даже |
при совместном |
||||||||||||||||||||||
какого-либо |
гена. |
|
|
|
|
|
|||||||||||||||||||
|
|
|
|
|
действии еще не могут обеспечить про- |
||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||
|
Теоретические расчеты показывают, |
текания |
эволюционного |
процесса. Для |
|||||||||||||||||||||
что влияние популяционных волн мо- |
этого нужны факторы, длительно дей- |
||||||||||||||||||||||||
жет быть особенно заметно в популя- |
ствующие в одном направлении. Один |
||||||||||||||||||||||||
циях очень малой величины обычно |
из них — изоляция. |
|
|
|
|
|
|
|
|||||||||||||||||
при |
численности |
|
размножающихся |
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||
|
|
|
|
|
|
|
|
|
i 22 Часть 3. Учение о м и к р о э в о л ю ц и и |
|
|
|
|
|
||||||||||
9.4. Изоляция как элементарный |
тельностью человека в биосфере все |
|||||||||||||||||||||||
|
|
эволюционный фактор |
|
чаще и чаще возникает подобная прост- |
||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
ранственная изоляция отдельных попу- |
|||||||||||
Изоляция |
— |
возникновение |
любых |
ляций внутри очень многих видов. Ти- |
||||||||||||||||||||
барьеров, |
|
ограничивающих |
|
|
панмиксию. |
пичным примером стало возникновение |
||||||||||||||||||
Значение изоляции в процессе эволю- |
в Евразии к началу XX в. разорван- |
|||||||||||||||||||||||
ции и сводится к нарушению свобод- |
ного |
ареала |
у |
соболя |
(Martes |
|
zibelli- |
|||||||||||||||||
ного скрещивания, что ведет к увели- |
па) — результат |
интенсивного промыс- |
||||||||||||||||||||||
чению и закреплению различий между |
ла (рис. 9.5). Обычно возникновение |
|||||||||||||||||||||||
популяциями |
и |
отдельными |
частями |
подобного разорванного ареала служит |
||||||||||||||||||||
всего населения вида. Без такого |
опасным симптомом возможного исчез- |
|||||||||||||||||||||||
закрепления |
эволюционных |
|
различий |
новения |
вида. |
|
|
|
|
|
||||||||||||||
невозможно |
никакое |
формообразова- |
|
Пространственная изоляция |
может |
|||||||||||||||||||
ние. |
|
|
|
|
|
|
|
|
|
|
|
|
возникнуть внутри видов малоподвиж- |
|||||||||||
Разнообразие |
форм |
и |
проявлений |
ных животных и растений, не разде- |
||||||||||||||||||||
изоляции в природе так велико, что для |
ленных заметными физико-географи- |
|||||||||||||||||||||||
понимания эволюционной роли изоля- |
ческими |
|
барьерами. |
Известно, |
что |
|||||||||||||||||||
ции необходимо кратко описать основ- |
обыкновенный соловей |
(Luscinia |
lusci- |
|||||||||||||||||||||
ные ее проявления в природе. |
|
nia), населяющий многие районы цент- |
||||||||||||||||||||||
Классификация |
явлений |
|
изоляции. |
ральной части европейской территории |
||||||||||||||||||||
В природе существуют: пространствен- |
СССР, в настоящее время практи- |
|||||||||||||||||||||||
ная |
и |
биологическая |
изоляции. |
|
чески |
сплошь |
находит |
подходящие |
||||||||||||||||
Пространственная изоляция |
может |
условия для гнездования как в необжи- |
||||||||||||||||||||||
существовать в разных формах: водные |
тых человеком местах, так и в зарослях |
|||||||||||||||||||||||
барьеры разделяют население «сухо- |
по обочинам дорог, в парках и даже |
|||||||||||||||||||||||
путных» видов, а барьеры суши изоли- |
скверах больших городов. При этом |
|||||||||||||||||||||||
руют население видов-гидробионтов, |
наблюдается четко выраженная кли- |
|||||||||||||||||||||||
возвышенности |
изолируют |
равнинные |
нальная1 |
|
изменчивость пения этих |
|||||||||||||||||||
популяции, а равнины — горные попу- |
птиц: по |
|
числу |
«коленец», по |
тембру |
|||||||||||||||||||
ляции и т. д. Сравнительно мало- |
и |
другим |
особенностям |
существуют |
||||||||||||||||||||
подвижные |
|
животные — наземные |
постепенные переходы от одной мест- |
|||||||||||||||||||||
моллюски на Гавайских островах, в |
ности |
к |
другой |
(при |
этом известно, |
|||||||||||||||||||
каждой из долин возникает самостоя- |
что характер песни наследственно обус- |
|||||||||||||||||||||||
тельная популяция со своими специ- |
ловлен) . Возникновение такой кли- |
|||||||||||||||||||||||
фическими |
особенностями. |
|
|
|
нальной изменчивости возможно толь- |
|||||||||||||||||||
Возникновение |
территориально- |
ко |
потому, |
что |
соловьи, |
несмотря на |
||||||||||||||||||
существование у них частых сезонных |
||||||||||||||||||||||||
механической |
изоляции |
объясняется |
||||||||||||||||||||||
перелетов, |
обладают большим |
гнездо- |
||||||||||||||||||||||
историей развития видов на определен- |
||||||||||||||||||||||||
вым консерватизмом: молодые |
возвра- |
|||||||||||||||||||||||
ных |
территориях (рис. |
9.4). В |
приве- |
|||||||||||||||||||||
щаются |
практически на |
то же |
место, |
|||||||||||||||||||||
денных |
случаях |
основной |
|
причиной |
||||||||||||||||||||
|
где |
они |
вывелись. |
|
|
|
|
|||||||||||||||||
изоляции |
стало |
наступление |
ледников. |
|
|
|
|
|||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||
За время, прошедшее после ледников, |
|
Рассмотрев |
пространственную |
изо- |
||||||||||||||||||||
изолированные формы еще не приоб- |
ляцию внутри вида, мы пришли к зак- |
|||||||||||||||||||||||
рели |
|
значительных |
морфофизиологи- |
лючению о существовании двух ее |
||||||||||||||||||||
ческих различий и относятся к единым |
проявлений: |
изоляция |
|
какими-либо |
||||||||||||||||||||
видам. Дальнейшие исследования могут |
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||
показать, |
что |
это формы |
уже |
возник- |
|
1 |
Клинальной изменчивостью принято назы- |
|||||||||||||||||
шего видового ранга. Полной изоляции |
|
|||||||||||||||||||||||
вать количественное варьирование признака или |
||||||||||||||||||||||||
на протяжении жизни десятков тысяч |
комплекса признаков популяций в определенной |
|||||||||||||||||||||||
поколений |
обычно |
бывает |
достаточно |
географической |
последовательности (Дж. Хакс- |
|||||||||||||||||||
для |
возникновения |
видов. |
|
|
|
|
ли) , осуществляемое в конечном итоге действием |
|||||||||||||||||
В |
настоящее |
время |
в |
связи |
с дея- |
естественного отбора в градиенте физико- |
||||||||||||||||||
географических |
условий. |
|
|
|
|
|||||||||||||||||||
Глава 9. Элементарные факторы эволюции |
135 |
Рис. 9.4. Примеры разорванных ареалов: ареалы голубой сороки (Cyanopica суапа) и вьюна (Misgurnus fossilis) (из Н. В. Тимофеева-Ресовского и др., 1977)
барьерами между частями видового на- |
(послекопуляционные). Первые меха- |
|||||||||||||||
селения |
и |
изоляция, |
определяемая |
низмы |
предотвращают |
потерю |
гамет, |
|||||||||
большей |
возможностью |
|
спаривания |
вторые связаны с потерей гамет и зигот |
||||||||||||
близко живущих особей, т. е. изоляция |
(Э. Майр). |
|
|
|
|
|||||||||||
расстоянием. |
|
|
|
|
|
|
Спариванию близких форм препят- |
|||||||||
Разбирая основные |
характеристики |
ствуют различия во время половой |
||||||||||||||
популяции, |
мы |
|
подчеркнули значение |
активности и созревания половых про- |
||||||||||||
радиуса |
репродуктивной |
|
активности |
дуктов. Известно существование «яро- |
||||||||||||
для |
особей |
вида |
(см. гл. |
7). Возник- |
вых» и «озимых» рас у миног (Lampet- |
|||||||||||
новение |
пространственной |
изоляции |
га) и некоторых лососевых рыб |
(Опсог- |
||||||||||||
связано с этой |
величиной. |
|
|
hynchus), |
которые резко |
отличаются |
||||||||||
Радиус |
репродуктивной |
активности |
временем нереста; между особями каж- |
|||||||||||||
наземных |
|
моллюсков |
обычно |
равен |
дой из рас существует высокая сте- |
|||||||||||
нескольким десяткам метров, а чир- |
пень изоляции. Среди растений извест- |
|||||||||||||||
ков — тысяче |
километров. |
Значение |
ны случаи генетически |
обусловленного |
||||||||||||
физико-географических барьеров в |
сдвига в период цветения, создающего |
|||||||||||||||
пространственной изоляции |
связано с |
биологическую изоляцию этих форм — |
||||||||||||||
биологическими |
|
особенностями |
вида в |
явление |
фенологического |
полиморфиз- |
||||||||||
целом. |
|
|
|
|
|
|
|
|
ма (рис. |
9.6). |
|
|
|
|
||
Биологическую |
изоляцию |
обеспечи- |
В |
природе |
обычна |
.биотопическая |
||||||||||
вают две группы механизмов: устра- |
изоляция, |
при |
которой |
потенциальные |
||||||||||||
няющие |
скрещивание |
(докопуляцион- |
партнеры по спариванию не встречают- |
|||||||||||||
ные) |
и |
изоляция при |
скрещивании |
ся, так как они чаще обитают в разных |
||||||||||||

Рис. 9.5. Пространственная структура ареала соболя (Martes zibellina) (по А. А. Насимовичу и В. В. Тимофееву, 1973):
1—границы ареала, 2—оставшиеся популяционные островки населения в 30-е годы XX в., 3—территория, занимаемая соболем в 70-е годы XX в.
местах. Так, часть зябликов (Fringilla coelebs) гнездится в Московской области в лесах таежного типа, а другая — в невысоких и редких насаждениях с большим числом полян. Потенциальная возможность перекрестного спаривания особей этих групп несколько ограничена. Интересный пример биотопической изоляции — симпатрические внутривидовые формы у обыкновенной кукушки (Cuculus с an or us).
В Европе обитает несколько «биологических рас» кукушек, различающихся генетически закрепленной окраской яиц. В Восточной Европе одни откладывают голубые яйца в гнезда обыкновенной горихвостки и лугового чекана, другие — светлые в крапинку яйца в гнезда мелких воробьиных птиц, имеющих яйца сходной окраски. Изоляция между этими формами кукушек поддерживается за счет уничтожения вида- ми-хозяевами недостаточно замаскированных яиц. У многих видов предпочтение биотопа — эффективный изоляционный механизм.
Большое значение в возникновении и поддержании биологической изоляции у близких форм имеет отологическая изоляция — осложнения спаривания, обусловленные особенностями поведения. Вскрыто большое разнообразие и распространение способов этологической изоляции у животных. Возможно, у животных этологические механизмы — наиболее обширная группа из докопуляционных изолирующих механизмов. Ничтожные на первый взгляд отличия в ритуале ухаживания и обмене зрительными, звуковыми, химическими раздражителями будут препятствовать продолжению ухаживания. Таков в общих чертах механизм этологической изоляции.
На рис. 9.7 показан характер световых вспышек у северо-американских светляков рода Photurus. Хорошо видно, что различие между близкими видами, а иногда и разными группами популяций внутри вида определяется длительностью, частотой и интенсивностью вспышек.
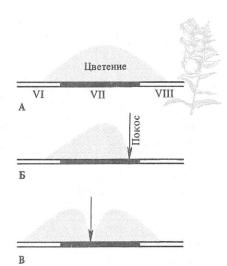
Рис. 9.6. На некошенных лугах большой погремок (Alectorolophus major) цветет и плодоносит в течение всего лета (А). При кошении в конце лета образовалась раса погремка, успевающая принести семена до начала покоса
(Б). При более раннем покосе |
(В) возникли |
две расы—ранневесенняя и |
позднеосенняя. |
Позднеосенняя раса содержала растения, замедленно развивающиеся до начала покоса, очень низкие, не попадавшие под косу, но затем быстро зацветающие и успевающие дать семена до начала морозов. Пример фенологического полиморфизма (ориг. рис. по данным Н. В. Цин-
гера, 1920)
Важным изолирующим механизмом, затрудняющим скрещивание близких видов, оказывается возникновение морфофизиологических различий в органах размножения (морфофизиологическая изоляция).
Развитие сложных по форме цветов у ряда насекомоопыляемых растений связано с приспособлением цветка к виду насекомых-опылителей.
У животных близких видов различия копулятивных органов особенно характерны для некоторых легочных моллюсков, насекомых, а среди млекопитающих — ряда групп грызунов.
Описанные механизмы изоляции обычно достаточны для предотвращения в природных условиях возможности скрещивания форм, принадлежащих к разным видам, и понижения эффективности скрещивания у внутривидо-

i 22 Часть 3. Учение о микроэволюции
Время, с
Вяд |
|
0 |
|
1 |
|
2 |
|
|
3 |
|
|
4 |
|
s |
|
|
|
|
|
|
|
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
cinctipennis |
|
|
|
|
|
|
|
|
|
|
|
I |
|
|
|
hebes |
|
: |
|
|
|
5 |
|
|
|
|
|
• |
|
|
|
potomaca |
1 |
i |
i |
|
§ |
! |
|
|
1 |
|
1 |
|
i |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
frontalis |
|
» |
|
i |
|
i |
|
|
8 |
|
|
1 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
versicolor, |
|
iilSIS |
|
|
|
|
|
|
|
|
|
|
i»SI |
|
|
разные подвиды |
|
pee |
|
|
|
|
|
|
|
|
|
|
|
!й |
|
|
|
Hi |
|
|
|
|
|
|
|
|
|
|
|
И |
|
fairehildi |
|
9 |
1 |
|
|
|
|
|
|
|
|
|
|
I |
|
lacicrescens |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
pensylvanica |
|
T33B |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
pyraiomimus |
|
m |
|
|
|
|
|
|
|
|
|
|
|
|
|
caerulucens |
|
ШЩ |
|
|
|
|
|
|
|
|
ШН |
|
|
|
|
iremaians |
|
•ЛПШШ |
|
|
|
|
|
|
|
|
|
|
|
||
|
|
ТппТПл |
|
|
|
|
|
|
|
|
|
|
|
||
Рис. 9.7. Пример эколого-этологической изоляции. Характеристика световых вспышек северо-
американских светляков рода |
Photurus. Высота |
и длина пятен указывает |
на интенсивность |
и длительность вспышки (из Э. Майра, 1968)
вых форм, далеко разошедшихся в про-
цессе эволюции (подвидов, |
групп уда- |
ленных географических |
популяций |
и т. п.). |
|
Вторая большая группа изолирующих механизмов в природе связана с возникновением изоляции после оплодотворения (собственно-генетическая изоляция), включающей гибель зигот после оплодотворения, развитие полностью или частично стерильных гибридов, а также пониженную жизнеспособность гибридов.
При межвидовом спаривании часто образуются вполне жизнеспособные гибриды, но у них, как правило, не развиваются нормальные половые клетки. В случае же нормального развития гамет гибриды оказываются малоплодовитыми. В природе есть случаи такой «изоляции посредством гибридизации»: на границе обитания двух близких форм постоянно существует зона, населенная вполне жизнеспособными гибридными особями, но их потомство либо ослаблено
и не выдерживает конкуренции с более сильными особями родительских видов, либо нежизнеспособно. На существование такого «гибридизационного» барьера между видами обычно указывает наличие в природе устойчивых гибридных зон между близкими видами. Такие гибридные зоны известны для некоторых насекомых, серой и черной ворон в Европе (рис. 9.8).
Значение изоляции в эволюции. Изоляция как эволюционный фактор не создает новых генотипов или внутривидовых форм. Значение изоляции в процессе эволюции состоит в том, что она закрепляет и усиливает начальные стадии генотипической дифференцировки, а также в том, что разделенные барьерами части популяции или вида неизбежно попадают под различное давление отбора. Изоляция ведет к сохранению специфичности генофонда дивергирующих форм.
Важная характеристика действия изоляции как фактора эволюции — это ее длительность. В большинстве случаев причина возникновения биологической или пространственной изоляции сохраняется на длительное время.
Действие изоляции на эволюционный материал по существу статистично и ненаправленно, и в этом изоляция сходна с другими эволюционными факторами (мутации и популяционные волны).
Существенный результат изоляции на микроэволюционном уровне — возникновение гомозиготизации на окраинах ареала.
Сравнивая относительное значение давления элементарных эволюционных факторов, надо отметить, что давление изоляции обычно превосходит давле-
ние мутационного |
процесса, и, видимо, |
|||
близко |
к величине |
давления |
волн |
жиз- |
ни (в |
конкретных |
условиях |
эти |
вели- |
чины могут быть |
различными). |
|
||
Изоляция расчленяет исходные популяции на две или более, а группы популяций — на различающиеся формы. Любая группа особей в природе

Глава 9. Элементарные факторы эволюции 139
Рис. 9.8 Пример изоляции посредством гибридизации: соприкосновение серой (1) (Corvus
согах) |
и черной |
(2) |
ворон |
(С. |
согопе). |
А—общий ареал; Б—фрагмент с |
изображением |
||||
зоны |
гибридизации |
(черная |
полоса) |
(по |
|
|
Э. Майру, |
1968). |
|
|
|
Гибриды оказываются менее жизнеспособными по сравнению с родительскими формами. Судя по неодинаковой ширине гибридной зоны (и некоторому колебанию ширины зоны в разные годы), относительная жизнеспособность гибридов различная в разных поколениях и частях ареала
давлением изоляции отделена от других близких групп.
Следовательно, |
действие |
изоля- |
ции — обязательное |
условие |
всякого |
достаточно длительного этапа эволюционного процесса.
Оценивая роль трех элементарных эволюционных факторов, можно сказать, что первые два фактора (мутационный процесс и волны жизни)— факторы-поставщики элементарного эволюционного материала (хотя и действуют совершенно по-разному), а изоляция оказывается факто- ром-усилителем генетических различий между группами особей.
Механизм действия трех факторо'в различен, но общим для них является ненаправленностъ, неопределенность и стохастичностъ действия. В природе три эволюционных фактора действуют совместно, однако роль каждого может усиливаться в зависимости от конкретной обстановки. Даже совместное действие трех элементарных факторов не обеспечивает устойчивого протекания направленного процесса эволюции.
Направленность эволюции придает действие естественного отбора.
Глава 10
Естественный отбор — движущая и направляющая сила эволюции
В предыдущей главе показано значение мутационного процесса, волн жизни и изоляции для эволюции. Эти элементарные факторы эволюции действуют ненаправленно. В то же время эволюция в целом — не хаотический, а направленный процесс, связанный с выработкой новых и новых приспособлений, возникновением одних и вымиранием других видов, возникновением иерархической системы таксонов, осуществлением прогрессивного развития живой природы. Известен лишь один направленный эволюционный фактор — естественный отбор.
В настоящее время учение о естественном отборе пополнено новыми фактами и развито множеством новых подходов. Однако все основные положения, развитые Ч. Дарвином в обосновании учения об отборе, полностью сохраняют свое значение.
10.1. Значение данным селекции для вскрытие механизма действия естественного отбора
Теория естественного отбора возникла на широком фундаменте практической селекции (см, гл. 3). На примере возникновения разных пород голу-
бей Ч. Дарвин убедительно показал, что все они получены человеком благодаря размножению тех особей, которые оказывались носителями каких-то желательных признаков или свойств. Таким же образом были созданы человеком все без исключения, порой причудливые, формы домашних животных и культурных растений, предназначен-
ные |
для удовлетворения тех или |
иных |
потребностей человека. |
Результаты искусственного отбора |
|
стали особенно ощутимыми с началом
систематического |
применения |
его в |
методической (сознательной) |
форме |
|
с XVIII в. При этом селекционер зара- |
||
нее определяет |
основные показатели |
|
будущей породы или сорта и старается затем реализовать свой идеал путем подбора родителей для скрещивания. Эффект такого отбора увеличивается при большой выборке исходных особей — это повышает вероятность быстрого нахождения особей с нужными уклонениями.
В бессознательном отборе Ч. Дарвин увидел аналогию процесса, действующего в природе независимо от человека. Только вместо человека, определяющего оставление для размножения нужных ему особей, в природе занимает процесс, действующий без