

Эволюционное учение
.pdf
|
|
|
Глава 10. Естественный отбор — д в и ж у щ а я и |
направляющая |
сила |
|
эволюции |
|
151 |
||||||||||||||||||||
реализации части потенциала размно- |
деляют возможность сохранения сход- |
||||||||||||||||||||||||||||
жения (см. гл. 9) образуют популяции, |
ных |
мутаций |
|
(в |
дальнейшем |
замена |
|||||||||||||||||||||||
устойчивые к |
действию |
ядов. |
|
|
фенокопий генокопиями). Отбор му- |
||||||||||||||||||||||||
|
У |
устойчивых |
|
форм |
обнаружен |
тантных особей ярового рыжика, слу- |
|||||||||||||||||||||||
ряд |
|
мутаций, |
определяющих |
синтез |
чайно оказавшихся сходными с льном, |
||||||||||||||||||||||||
фермента ацетилхолинэстеразы, проис- |
привел к формированию нового вида |
||||||||||||||||||||||||||||
ходит отбор на фиксацию в популя- |
сорняка — льняного рыжика. |
|
|
||||||||||||||||||||||||||
ции |
алеллей, вызывающих разрушение |
|
Другой |
|
пример — формирование |
||||||||||||||||||||||||
инсектицида в организме, по формиро- |
фенологического |
полиморфизма |
у |
||||||||||||||||||||||||||
ванию структур покровов, ограничи- |
погремка |
|
Rhinanthus |
|
(Alectorolophus) |
||||||||||||||||||||||||
вающих |
поступление |
|
яда |
в организм, |
под влиянием сроков и числа покосов. |
||||||||||||||||||||||||
и по формированию поведения, связан- |
Как показал Н. В. Цингер, R. major |
||||||||||||||||||||||||||||
ного с избеганием контактов с пести- |
представляет собой комплекс форм, от- |
||||||||||||||||||||||||||||
цидом. Так, например, у комнатной |
личающихся друг от друга продол- |
||||||||||||||||||||||||||||
мухи за 30 лет было обнаружено 9 раз- |
жительностью жизни и морфологичес- |
||||||||||||||||||||||||||||
ных |
мутаций |
(в |
разных |
хромосомах), |
ки. На нескашиваемых участках встре- |
||||||||||||||||||||||||
определяющих |
устойчивость к |
разным |
чается форма, цветущая в течение |
||||||||||||||||||||||||||
инсектицидам. |
|
|
|
|
|
|
|
|
|
июня — августа |
(R. т. |
|
montanus). |
На |
|||||||||||||||
|
Эти |
явления |
известны и при повы- |
участках, скашиваемых в июне, у боль- |
|||||||||||||||||||||||||
|
шого |
|
погремка |
дифференцировалась |
|||||||||||||||||||||||||
шении |
устойчивости |
микроорганизмов |
|
||||||||||||||||||||||||||
раннеспелая раса, |
тогда |
как |
на |
участ- |
|||||||||||||||||||||||||
к антибиотикам |
и |
другим |
лекарствен- |
||||||||||||||||||||||||||
ках, скащиваемых в июле,— позднеспе- |
|||||||||||||||||||||||||||||
ным |
препаратам. |
|
|
|
|
|
|
|
|||||||||||||||||||||
|
|
|
|
|
|
|
лая. На пашне и в посевах обнаружена |
||||||||||||||||||||||
|
Ч. |
Дарвин часто |
расшифровывал |
||||||||||||||||||||||||||
|
сорная |
форма (R. |
т. |
apters), которая |
|||||||||||||||||||||||||
творческую роль |
естественного |
отбора |
|||||||||||||||||||||||||||
отличается |
от |
ранее |
упомянутых |
не |
|||||||||||||||||||||||||
путем сравнения |
его |
|
с |
искусственным |
|||||||||||||||||||||||||
|
только морфологически, но и по срокам |
||||||||||||||||||||||||||||
отбором. В этой |
связи |
интересны два |
|||||||||||||||||||||||||||
цветения |
|
и |
созревания, |
неосыпае- |
|||||||||||||||||||||||||
примера. |
Первый |
— |
исследование |
|
|||||||||||||||||||||||||
мостью |
семян |
и |
т. |
д. |
|
Аналогичный |
|||||||||||||||||||||||
Н. |
В. |
Цингером |
льняного |
рыжика |
|
||||||||||||||||||||||||
механизм |
|
предполагается |
и |
для |
обра- |
||||||||||||||||||||||||
(Camelina |
linicola) — |
|
сорняка в посевах |
|
|||||||||||||||||||||||||
|
зования |
подвидов |
R. |
|
т. |
vernalis |
и |
||||||||||||||||||||||
льна. Анализ показал, что сорняк обра- |
|
||||||||||||||||||||||||||||
R. |
т. |
aestivalis, |
описываемых некото- |
||||||||||||||||||||||||||
зовался под действием бессознательно- |
|||||||||||||||||||||||||||||
рыми авторами как виды. В рассматри- |
|||||||||||||||||||||||||||||
го |
и |
естественного |
отбора из |
ярового |
|||||||||||||||||||||||||
ваемых |
случаях |
приспособления яви- |
|||||||||||||||||||||||||||
рыжика |
|
(С. |
glabrata), |
разводимого |
в |
||||||||||||||||||||||||
|
лись |
следствием |
отбора |
особей |
по |
||||||||||||||||||||||||
культуре для получения масла. Семена |
|||||||||||||||||||||||||||||
индивидуальным |
особенностям |
(см. |
|||||||||||||||||||||||||||
ярового рыжика, как и любого другого |
|||||||||||||||||||||||||||||
рис. |
9.6). |
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||
растения, |
обнаруживают |
наследствен- |
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||
ные различия по величине, массе, цвету, |
|
Рассмотренные |
примеры |
действия |
|||||||||||||||||||||||||
темпам прорастания и т. д. Семена яро- |
отбора в природе и эксперименте сви- |
||||||||||||||||||||||||||||
вого рыжика заносились в посевы льна |
детельствуют |
о |
существовании |
отбора |
|||||||||||||||||||||||||
человеком или естественным путем. По |
и его решающей роли в создании и |
||||||||||||||||||||||||||||
внешнему виду льняной рыжик сходен |
распространении различных |
свойств |
и |
||||||||||||||||||||||||||
с льном: тонкий и мягкий стебель, |
признаков, |
вплоть |
до |
возникновения |
|||||||||||||||||||||||||
узкие и тонкие листья, вытянутые меж- |
новых |
видов. |
|
|
|
|
|
|
|
|
|
||||||||||||||||||
доузлия. Опыты показали, что при вы- |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||
ращивании ярового рыжика в густых и |
|
10.6. Эффективность и скорость |
|||||||||||||||||||||||||||
увлажненных посевах льна уже в пер- |
|
||||||||||||||||||||||||||||
|
действия |
естественного |
©тборз |
|
|||||||||||||||||||||||||
вом поколении они приобретают осо- |
|
|
|||||||||||||||||||||||||||
бенности, |
свойственные |
льняному ры- |
|
Эффективность и скорость действия |
|||||||||||||||||||||||||
жику |
(увеличивается |
высота стебля; |
в |
|
|||||||||||||||||||||||||
естественного |
отбора |
зависят от ряда |
|||||||||||||||||||||||||||
посевах льна высокие стебли позволяют |
|||||||||||||||||||||||||||||
факторов: условий существования, кон- |
|||||||||||||||||||||||||||||
избежать недостатка света). Эти изме- |
|||||||||||||||||||||||||||||
кретных |
признаков и |
в |
первую оче- |
||||||||||||||||||||||||||
нения хотя и не наследуются, но опре- |
|||||||||||||||||||||||||||||
редь — от |
величины |
давления |
отбора |
||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||
(т. е. степени количественного воз- |
ром —0,01. |
В |
природных |
|
условиях |
||||||||||||||||
действия) . Для оценки величины давле- |
коэффициент отбора обычно не превы- |
||||||||||||||||||||
ния отбора необходимо познакомить- |
шает 0,10—0,20, чаще имеет меньшие |
||||||||||||||||||||
ся с понятием адаптивная ценность |
значения. Конечно, по отношению к |
||||||||||||||||||||
генотипа. |
|
|
|
|
|
|
|
|
летальным мутациям, резко понижаю- |
||||||||||||
В |
процессе отбора индивидуальные |
щим |
жизнеспособность |
организмов, |
|||||||||||||||||
достоинства каждого генотипа в попу- |
коэффициент отбора может быть и |
||||||||||||||||||||
ляциях |
оцениваются |
по |
способности |
очень значителен или достигает 1,0. |
|||||||||||||||||
передавать гены будущим поколениям. |
Коэффициент отбора может выра- |
||||||||||||||||||||
Способность генотипа к выживанию и |
жать как возрастание, так и убывание |
||||||||||||||||||||
воспроизведению |
по |
сравнению |
с |
дру- |
концентрации аллеля. В первом случае |
||||||||||||||||
гими генотипами в популяциях называ- |
имеет |
место |
размножение |
носителя, |
|||||||||||||||||
ется |
адаптивной |
ценностью генотипа. |
во втором — его элиминация. |
|
|
||||||||||||||||
Она характеризует степень относи- |
В природе оценка адаптивной цен- |
||||||||||||||||||||
тельной приспособленности генотипа и |
ности генотипа по отдельным призна- |
||||||||||||||||||||
обозначается обычно буквой «W», ко- |
кам происходит редко, обычно отбор |
||||||||||||||||||||
торая колеблется в пределах от 0 до 1. |
идет по комплексу признаков. Для |
||||||||||||||||||||
При |
«W»= 0 |
передача |
генетической |
характеристики |
адаптивной |
ценности |
|||||||||||||||
информации |
индивидуума следующему |
всех генотипов в популяции опреде- |
|||||||||||||||||||
поколению отсутствует и аллель исче- |
ляют |
среднюю |
приспособленность |
W. |
|||||||||||||||||
зает |
из |
популяции; |
при |
«W»= 1 |
воз- |
Она равна сумме частот генотипов |
|||||||||||||||
можно |
образование |
максимального |
после отбора и определяется по фор- |
||||||||||||||||||
числа гамет с данным наследственным |
муле |
Харди—Вайнберга |
(см. |
гл. |
8): |
||||||||||||||||
признаком |
(полностью |
реализуются |
|
|
|
|
|
|
|
|
|
|
|||||||||
потенциальные возможности к размно- |
|
W=p2W0+2pqW\+q2W2. |
|
|
|
||||||||||||||||
жению) . |
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
Адаптивная |
ценность линотипа |
оп- |
При этом среднюю приспособлен- |
||||||||||||||||||
ределяется |
всем |
комплексом |
генов, |
ность доминантной гомозиготы при- |
|||||||||||||||||
поэтому даже генотипы, |
гомозиготные |
нимают за 1, а приспособленность |
|||||||||||||||||||
по одному и тому же гену, могут |
остальных |
генотипов |
соотносят к |
ней |
|||||||||||||||||
обладать различной адаптивной цен- |
в процентах. |
|
|
|
|
|
|
|
|||||||||||||
ностью. |
Количественно |
|
адаптивную |
Генотипы |
|
Средняя |
приспособленность |
||||||||||||||
ценность |
выражают в % |
частот алле- |
|
АА |
|
|
|
W0=W0/W0=l |
|
|
|||||||||||
лей до и после отбора, |
т. |
е. |
учетом |
|
|
|
|
|
|
||||||||||||
|
Аа |
|
|
W\ = Wx/W0=l—S |
|
||||||||||||||||
их частот в поколениях, что в свою |
|
аа |
|
|
Wi=W2/W0=\— S2 |
|
|||||||||||||||
очередь зависит от коэффициента отбо- |
|
|
|
|
|
|
|
|
|
|
|||||||||||
ра. |
|
|
|
|
|
|
|
|
|
|
Применяя эти формулы и зная |
||||||||||
Коэффициент |
отбора |
характеризует |
соотношение генотипов до и после |
||||||||||||||||||
интенсивность элиминации или сниже- |
отбора, легко определить коэффици- |
||||||||||||||||||||
ние воспроизведения мутантного алле- |
ент отбора (табл. 10.3). |
|
|
|
|
||||||||||||||||
ля по сравнению с исходной формой. |
Чем |
значительнее |
коэффициент |
||||||||||||||||||
Обозначается он буквой «5» и представ- |
отбора, тем интенсивнее отбор против |
||||||||||||||||||||
ляет |
величину, |
обратную |
адаптивной |
каких-либо генотипов, т. е. выше дав- |
|||||||||||||||||
ценности генотипа: варьирует от 1 до 0. |
ление отбора. Давление отбора за от- |
||||||||||||||||||||
Чем больше адаптивная ценность гено- |
резок времени всегда приводит к до- |
||||||||||||||||||||
типа, тем ниже коэффициент отбора. |
стижению |
|
определенного |
|
положи- |
||||||||||||||||
Если W— 1, то 5 = 0 . Если из 100 родив- |
тельного результата (изменению ча- |
||||||||||||||||||||
шихся особей с определенным призна- |
стот генов). Это эффективность отбо- |
||||||||||||||||||||
ком до размножения доживут все, а с |
ра. |
|
|
|
|
|
|
|
|
|
|||||||||||
другим признаком — лишь |
99 |
особей, |
Отбор |
особенно |
эффективен |
про- |
|||||||||||||||
то можно сказать, что в первом случае |
|||||||||||||||||||||
тив доминантных мутаций и при усло- |
|||||||||||||||||||||
коэффициент отбора равен 0, а во вто- |
|||||||||||||||||||||
вии их полного |
выпадения |
и |
проявле- |
||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|||||||||||
Глава 10. Естественный отбор — движущая и направляющая сила эволюции |
153 |
Рис. 10.6. Отбор против доминантного (пунк- |
Рис. 10.7. Отбор против доминантных (//) и |
||||
тиром) |
и |
рецессивного |
генотипа при |
давлении |
рецессивных (/) признаков при низких и высоких |
отбора |
5 = 1 . Исходная |
концентрация |
генотипов |
концентрациях (по В. Людвигу, 1940) |
|
равна |
1% |
(из Н. П. Дубинина, 1970) |
|
|
|
ния. При 5 = 1 популяция за одно поколение избавляется от доминантных летальных мутаций (рис. 10.6). Например, отсутствие насыщения попу-" ляций человека мутацией одной из форм карликовости (ахондроплазия) объясняется ее доминантностью и устранением в результате отбора. Особи, несущие ее, умирают в раннем возрасте, и по этой причине она не накапливается в популяциях человека. Отбор против рецессивных генов и при неполном доминировании, наоборот, затруднен. В гетерозиготном состоянии (под «прикрытием» нормального
фенотипа) они как бы ускользают от действия отбора. Особенно трудно избавить от рецессивных мутаций большие популяции, где их перевод в гомозиготное состояние происходит очень медленно (рис. 10.7).
Эффективность отбора, как следует из данных рис. 10.7, в значительной степени зависит от исходной концентрации гена в популяции. При очень низких (/) и очень высоких (//) концентрациях отбираемого признака (гена) отбор действует слишком медленно, при средних концентрациях — очень быстро даже при низких вели-
Т а б л и ц а 10.3. Определение |
величины коэффициента отбора (5) генотипов |
|||
|
|
|
|
|
Показатели |
|
|
Генотипы |
|
|
АА |
|
Аа |
аа |
Частота до отбора |
0,25 |
|
0,50 |
0,25 |
|
|
|
|
|
Частота после |
0,35 |
|
0,48 |
0,17 |
отбора |
|
|
|
|
|
|
|
|
|
Относительная |
^0=0,35/0,25=1,4; |
|
ИЛ = 0,48/0,50= 0,96 |
1^2=0,17/0,25=0,68 |
выживаемость |
£ " = 1 , 4 / 1 , 4 = 1,0 |
|
W1 |
Wо |
|
|
— =0,96/1,4=0,7 |
— = 0,68/1,4 = 0,4 |
|
|
W о |
|
W о |
W о |
Коэффициент от- |
1,0—1,0=0 |
|
1,0—0,7=0,3 |
1,0—0,4=0,6 |
бора |
|
|
|
|
|
|
|
|
|
i 22 |
Часть 3. Учение о микроэволюции |
|
чинах коэффициента отбора. Расчеты показывают, что при величине коэффициента отбора, равной всего 0,01, в популяции с доминантным геном А и рецессивным а возрастание частоты первого гена пойдет следующим образом:
Возрастание, % |
Число поколений |
|
0,1 — 1 |
250 |
|
1 |
—50 |
500 |
50 |
—98 |
5000 |
98 |
—99 |
5000 |
99 |
—99,9 |
90 000 |
нака, устраняя от размножения все особи, фенотипически заметно уклоняющиеся в ту или другую сторону от сложившейся нормы (рис. 10.8).
Действие стабилизирующего отбора можно пояснить на многих примерах. Так, после снегопада и сильных ветров в Северной Америке было найдено 136 оглушенных и полуживых домовых воробьев (Passer domesticus);
72 из них выжили, а 64 погибли. У погибших птиц были очень длинные или очень короткие крылья. Особи со средними — «нормальными» — крыльями оказались более выносливыми.
|
|
|
|
|
|
|
|
|
|
|
Другой |
пример — размеры |
и |
фор- |
|||||||||
|
|
|
|
|
|
|
|
|
|
|
ма цветов у энтомофильных растений. |
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
Исследование ветро- и насекомоопы- |
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
ляемых растений показало, что раз- |
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
меры и форма цветов у энтомофиль- |
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
ных |
растений |
|
более |
стабильны, |
чем |
|||||||
|
|
|
|
|
|
|
|
|
|
|
у анемофильных. Устойчивость цвет- |
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
ков энтомофилов обусловлена сопря- |
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
женной эволюцией растений и их опы- |
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
лителей, |
«выбраковкой» |
уклонившихся |
||||||||||
|
|
|
|
|
|
|
|
|
|
|
форм. Шмель |
|
не |
может |
проникнуть |
||||||||
|
|
|
|
|
|
|
|
|
|
|
в слишком узкий венчик цветка, хобо- |
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
ток бабочки не может коснуться слиш- |
||||||||||||
10.7. Основные формы естественного |
ком |
коротких |
тычинок |
у |
растений с |
||||||||||||||||||
длинным |
венчиком. |
|
Только растения |
||||||||||||||||||||
|
|
|
отбора |
|
|
|
|
|
|||||||||||||||
|
|
|
|
|
|
|
с цветками, строго |
соответствующими |
|||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||
В |
современной |
эволюционной тео- |
«стандартам», |
выработанным |
в |
тече- |
|||||||||||||||||
ние |
миллионов |
|
лет |
совместной |
эволю- |
||||||||||||||||||
рии |
вопрос |
о |
формах |
естественного |
|
||||||||||||||||||
цией цветковых растений и насекомых- |
|||||||||||||||||||||||
отбора |
остается |
одним из |
дискусси- |
||||||||||||||||||||
опылителей, могут оставить потомство. |
|||||||||||||||||||||||
онных. |
Выделяется |
более 30 различ- |
|||||||||||||||||||||
Известно, что обычно наибольшее чис- |
|||||||||||||||||||||||
ных |
форм |
отбора. |
Однако |
основных |
|||||||||||||||||||
ло особей |
в каждой |
популяции |
обла- |
||||||||||||||||||||
форм |
|
отбора |
в |
популяциях |
только |
||||||||||||||||||
|
дает |
средней |
выраженностью |
любого |
|||||||||||||||||||
три: |
стабилизирующий, |
движущий |
и |
||||||||||||||||||||
признака. Этот |
|
факт, |
подтвержденный |
||||||||||||||||||||
дизруптивный. |
|
|
|
|
|
|
|
|
|||||||||||||||
|
|
|
отбор — форма |
многими |
тысячами |
исследований |
раз- |
||||||||||||||||
Стабилизирующий |
|||||||||||||||||||||||
нообразных |
признаков, |
служит |
кос- |
||||||||||||||||||||
естественного |
|
отбора, |
направленная |
||||||||||||||||||||
|
венным |
доказательством |
всеобщности |
||||||||||||||||||||
на поддержание |
|
и |
повышение |
устой- |
|||||||||||||||||||
|
действия |
|
стабилизирующей |
формы |
|||||||||||||||||||
чивости |
реализации |
в |
популяции |
сред- |
|
||||||||||||||||||
отбора. |
|
|
|
|
|
|
|
|
|
|
|||||||||||||
него, |
|
ранее |
|
сложившегося |
значения |
|
|
|
|
|
|
|
|
|
|
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||
признака или свойства. При стабили- |
Один из результатов действия ста- |
||||||||||||||||||||||
зирующем |
отборе |
преимущество |
в |
билизирующего |
отбора — биохими- |
||||||||||||||||||
размножении получают особи со сред- |
ческое единство |
(«биохимическая уни- |
|||||||||||||||||||||
ним выражением признака (такой от- |
версальность») жизни на Земле. Так. |
||||||||||||||||||||||
бор поэтому образно называют «вы- |
аминокислотный состав низших по- |
||||||||||||||||||||||
живанием заурядностей»). Эта форма |
звоночных |
и человека |
почти |
один и |
|||||||||||||||||||
отбора как бы охраняет и усиливает |
тот же, почти одинаков состав |
фер- |
|||||||||||||||||||||
установившуюся |
характеристику |
приз- |
ментов у разных растений и т. д. |
Био- |
|||||||||||||||||||
Глава 10. Естественный отбор — движущая и направляющая сила эволюции |
155 |
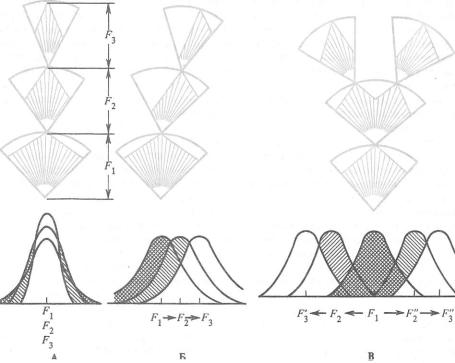
Рис. 10.8. Схема действия стабилизирующей ( А ) , движущей (Б) и дизруптивной (В) форм естественного отбора (по Н. В. Тимофееву-Ресовскому и др., 1977).
F—поколения. На популяционных кривых заштрихованы элиминируемые варианты. Величина дуги при отборе внутри одного потомства соответствует норме реакции
химические основы жизни, |
возникшие |
ния мутационного процесса, выбра- |
||||||||||
на заре биологической эволюции, ока- |
ковывая уклонения от приспособи- |
|||||||||||
зались надежными для воспроизведе- |
тельной нормы. Без него не было бы |
|||||||||||
ния организмов независимо от уровня |
устойчивости в живой природе. В этом |
|||||||||||
их организации. Об эволюции хими- |
состоит |
важная |
консервативная |
роль |
||||||||
ческого |
состава |
живых |
организмов |
естественного |
отбора, |
|
необходимая |
|||||
нередко говорят как о «теме с неболь- |
также для сохранения и усиления ре- |
|||||||||||
шими вариациями», что порой даже |
зультатов |
движущей |
формы отбора. |
|||||||||
используется для доказательства от- |
Необходимо |
еще раз |
подчеркнуть |
|||||||||
сутствия отбора вообще. На самом |
два аспекта действия стабилизирую- |
|||||||||||
деле в этом проявляется действие ста- |
щего отбора. С одной стороны, нега- |
|||||||||||
билизирующего отбора. |
|
|
тивный аспект, т. е. элиминацию всех |
|||||||||
Стабилизирующая форма |
отбора |
особей, отклоняющихся от «стандарт- |
||||||||||
в течение сотен тысяч и миллионов |
ного» фенотипа; иногда эта форма от- |
|||||||||||
поколений оберегает виды от сущест- |
бора определяется |
как |
нормализующий |
|||||||||
венных изменений. Она действует до |
отбор. С другой стороны, можно выде- |
|||||||||||
тех пор, пока условия жизни, при ко- |
лять и позитивный аспект стабилизи- |
|||||||||||
торых данный признак или свойство |
рующего отбора, который часто обо- |
|||||||||||
выработано, существенно не меняются. |
значается |
термином |
канализирующий |
|||||||||
Стабилизирующая форма отбора обе- |
отбор |
(К. |
Уоддингтон) — отбор |
ин- |
||||||||
регает |
норму от |
разрушающего |
влия- |
дивидуумов |
с |
генами, |
способными |
|||||
стабилизировать |
процесс |
онтогенеза |
и |
линии снизил их число с 32 до 25, а в |
|||||||||||||||||||||||
снижать его чувствительность ко вся- |
другой (после 20 поколений) повысил |
||||||||||||||||||||||||||
ким помехам |
(внутренним и внешним). |
до |
45—50 |
шт. (см. рис. 9.1). Генети- |
|||||||||||||||||||||||
В популяциях могут появляться гены, |
ческий анализ мух новых линий пока- |
||||||||||||||||||||||||||
понижающие |
жизнеспособность |
фено- |
зал |
наследственную |
обусловленность |
||||||||||||||||||||||
типа. В таких случаях отбор будет |
вновь возникших особенностей. В экс- |
||||||||||||||||||||||||||
направлен в пользу особей с комплек- |
периментах с тем же видом дрозофил |
||||||||||||||||||||||||||
сом генов (супергенов), обеспечиваю- |
показана возможность заметного по- |
||||||||||||||||||||||||||
щих устойчивый ход онтогенеза и спо- |
вышения |
|
холодоустойчивости |
после |
|||||||||||||||||||||||
собность к модификациям. Какая из |
отбора. Так, у исходной линии при |
||||||||||||||||||||||||||
двух |
форм |
стабилизирующего |
отбора |
температуре 0°С выживало лишь 28% |
|||||||||||||||||||||||
(нормализующий |
или |
|
канализирую- |
четырехдневных |
личинок. |
|
После |
30 |
|||||||||||||||||||
щий) |
окажется важнее, зависит от ин- |
поколений отбора для личинок темпе- |
|||||||||||||||||||||||||
тенсивности притока новых генов в |
ратура 0°С превратилась в фактор, |
||||||||||||||||||||||||||
популяцию (Э. Майр). |
|
|
|
|
|
стимулирующий развитие. Этот про- |
|||||||||||||||||||||
|
Движущий |
отбор. |
Движущей |
(или |
цесс характерен в природе; |
часто пер- |
|||||||||||||||||||||
|
воначально |
вредный |
фактор при |
его |
|||||||||||||||||||||||
направленной) |
|
формой |
отбора принято |
||||||||||||||||||||||||
|
постоянном |
действии |
превращается в |
||||||||||||||||||||||||
называть |
отбор, |
способствующий |
сдви- |
||||||||||||||||||||||||
фактор |
нормального |
развития. Анализ |
|||||||||||||||||||||||||
гу |
среднего |
|
значения |
|
признака |
или |
|||||||||||||||||||||
|
|
показал, |
|
что |
холодоустойчивость |
в |
|||||||||||||||||||||
свойства. |
Такой |
отбор |
способствует |
|
|||||||||||||||||||||||
эксперименте |
возникла |
не |
в |
результа- |
|||||||||||||||||||||||
закреплению новой нормы взамен ста- |
|||||||||||||||||||||||||||
те |
распространения |
единичной |
мута- |
||||||||||||||||||||||||
рой, |
пришедшей |
в |
несоответствие |
с |
|||||||||||||||||||||||
ции, вызванной действием |
холода, |
а в |
|||||||||||||||||||||||||
условиями. |
Изменение |
|
признака |
при |
|||||||||||||||||||||||
|
результате |
|
действия |
движущего |
отбо- |
||||||||||||||||||||||
этом может происходить как в сторону |
|
||||||||||||||||||||||||||
ра в пределах гетерогенной популяции. |
|||||||||||||||||||||||||||
усиления, так и ослабления. Боль- |
|||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||
шинство приводившихся ранее прямых |
|
Дизруптивный |
отбор. |
Эта |
форма |
||||||||||||||||||||||
и косвенных доказательств естествен- |
отбора существует в случаях, когда ни |
||||||||||||||||||||||||||
ного отбора относится именно к дви- |
одна из групп генотипов не получает |
||||||||||||||||||||||||||
жущей форме, отражающей класси- |
абсолютного |
преимущества |
в |
борьбе |
|||||||||||||||||||||||
ческое, дарвиновское, понимание сущ- |
за существование из-за разнообразия |
||||||||||||||||||||||||||
ности |
отбора. |
|
|
|
|
|
|
|
|
условий, |
одновременно |
встречающих- |
|||||||||||||||
|
Утрата |
признака — обычно |
ре- |
ся на одной территории. При этом в |
|||||||||||||||||||||||
зультат |
действия |
движущей |
формы |
одних условиях отбирается одно ка- |
|||||||||||||||||||||||
отбора. Например, в условиях функ- |
чество |
признака, |
в |
других — другое. |
|||||||||||||||||||||||
циональной непригодности органа |
(или |
Дизруптивный отбор направлен против |
|||||||||||||||||||||||||
его части) естественный отбор спо- |
особей со средним и промежуточным |
||||||||||||||||||||||||||
собствует их редукции. Утрата крыль- |
характером признаков и ведет к уста- |
||||||||||||||||||||||||||
ев у части птиц и насекомых, пальцев |
новлению |
полиморфизма |
в |
пределах |
|||||||||||||||||||||||
у копытных, конечностей у змей, глаз |
популяций. Популяция как бы «раз- |
||||||||||||||||||||||||||
у пещерных животных, корней и листь- |
рывается» по данному признаку на |
||||||||||||||||||||||||||
ев |
у |
растений-паразитов — примеры |
несколько групп. Поэтому дизруптив- |
||||||||||||||||||||||||
действия движущего отбора в направ- |
ный отбор называется также разры- |
||||||||||||||||||||||||||
лении редукции органов. Материал для |
вающим |
|
или |
расчленяющим. |
Итак, |
||||||||||||||||||||||
действия отбора в направлении редук- |
дизруптивной |
называется |
форма |
от- |
|||||||||||||||||||||||
ции органов доставляется разного рода |
бора, |
благоприятствующая |
|
более |
чем |
||||||||||||||||||||||
мутациями, которые ведут к дезин- |
одному фенотипу и действующая про- |
||||||||||||||||||||||||||
теграции организма и нарушению сис- |
тив |
средних |
промежуточных |
форм. |
|||||||||||||||||||||||
темы его корреляций (см. гл. 14). |
|
|
Во |
|
всех |
случаях |
полиморфизма |
||||||||||||||||||||
|
Была изучена возможность изме- |
действовал отбор дизруптивной фор- |
|||||||||||||||||||||||||
нения числа щетинок у дрозофилы |
мы. Рассмотрим пример действия та- |
||||||||||||||||||||||||||
(Drosophila melanogaster). Направлен- |
кого отбора по закреплению полимор- |
||||||||||||||||||||||||||
ный отбор через 30 поколений в одной |
физма |
по |
|
окраске раковин |
у |
земля- |
|||||||||||||||||||||
|
|
Глава |
10. |
Естественный отбор — движущая |
и направляющая сила эволюции |
|
|
157 |
||||||||||||||||||
ной улитки Cepaea nemoralis. Эти улит- |
|
Когда в |
лабораторной |
|
популяции |
|||||||||||||||||||||
ки различаются по |
количеству полос |
дрозофил менялось соотношение сам- |
||||||||||||||||||||||||
на раковине и степени пигментиро- |
цов, ведущих свое происхождение из |
|||||||||||||||||||||||||
ванное™ входного отверстия раковины |
двух |
|
разных |
природных |
популяций, |
|||||||||||||||||||||
в зависимости |
от |
окраски |
фона. Так, |
самцы, |
оказавшиеся |
в |
меньшинстве, |
|||||||||||||||||||
в лесах, где почвы коричневого цвета, |
участвовали в спариваниях непропор- |
|||||||||||||||||||||||||
чаще встречаются особи с коричневой |
ционально чаще самцов, составляю- |
|||||||||||||||||||||||||
и розовой окраской раковин, на участ- |
щих большинство. Во всех подобных |
|||||||||||||||||||||||||
ках с грубой и желтой травой преоб- |
случаях |
приспособленность |
генотипа |
|||||||||||||||||||||||
ладает желтая окраска и т. п. Подоб- |
(т. е. положительное давление отбо- |
|||||||||||||||||||||||||
ные различия в окраске раковин явно |
ра) |
будет |
функцией |
|
частоты |
этого |
||||||||||||||||||||
приспособительны, так как предох- |
генотипа. |
|
|
|
|
|
|
|
|
|
||||||||||||||||
раняют животных от истребления пти- |
|
Другая |
большая группа |
|
примеров |
|||||||||||||||||||||
цами. Полиморфизм улиток по окрас- |
действия естественного отбора связа- |
|||||||||||||||||||||||||
ке — результат |
действия |
|
дизруптив- |
на с выбором одной из двух главных |
||||||||||||||||||||||
ного отбора против средней нормы: |
стратегий размножения: либо корот- |
|||||||||||||||||||||||||
единая |
популяция |
распадается |
на |
кая продолжительность жизни, ранняя |
||||||||||||||||||||||
несколько форм и ни одна из них не |
половая зрелость и огромное число |
|||||||||||||||||||||||||
имеет решающего преимущества перед |
потомков, |
либо |
длительная |
жизнь |
||||||||||||||||||||||
другой. . |
|
|
|
|
|
|
|
|
|
особи, |
позднее |
наступление |
зрелости, |
|||||||||||||
Общий |
|
результат |
дизруптивного |
небольшое число потомков. И в первом |
||||||||||||||||||||||
отбора — формирование |
|
гетерозигот- |
(r-стратегия и соответственно г-отбор), |
|||||||||||||||||||||||
ного полиморфизма. Результат диз- |
и |
во |
|
втором |
случае |
(^-стратегия |
и |
|||||||||||||||||||
руптивного |
отбора |
(и |
возникновение |
соответственно АГ-отбор) достигается в |
||||||||||||||||||||||
его) можно представить в виде сов- |
конечном итоге сохранение популяции. |
|||||||||||||||||||||||||
местного |
и |
попеременного |
действия |
При r-отборе большая часть энергети- |
||||||||||||||||||||||
ведущего |
и |
стабилизирующего отбора. |
ческих затрат вида идет на появление |
|||||||||||||||||||||||
Другие |
|
формы |
|
естественного |
от- |
сравнительно многочисленного |
потом- |
|||||||||||||||||||
|
|
ства |
(обычно, |
в |
нестабильных |
усло- |
||||||||||||||||||||
бора. |
Как |
уже |
говорилось, |
к настоя- |
||||||||||||||||||||||
виях) , |
при |
.К-отборе — на |
выживание |
|||||||||||||||||||||||
щему |
|
времени |
выделено |
|
несколько |
|||||||||||||||||||||
|
|
исходно |
немногочисленного |
|
потомства |
|||||||||||||||||||||
десятков |
разных |
форм |
естественного |
|
||||||||||||||||||||||
(обычно |
в |
стабильных |
условиях сре- |
|||||||||||||||||||||||
отбора, |
связанных |
|
с отдельными |
спе- |
||||||||||||||||||||||
|
ды). |
|
Рассмотренные |
формы |
отбора |
|||||||||||||||||||||
цифическими сторонами действия еди- |
|
|||||||||||||||||||||||||
могут |
|
быть |
названы |
в |
общей |
форме |
||||||||||||||||||||
ного, |
|
всеобщего |
|
(«дарвиновского») |
|
|||||||||||||||||||||
|
|
плотностно-зависимым |
|
отбором |
(т. е. |
|||||||||||||||||||||
естественного отбора. Рассмотрим |
не- |
|
||||||||||||||||||||||||
отбором, |
связанным с |
плотностью |
на- |
|||||||||||||||||||||||
которые из этих специфических форм. |
||||||||||||||||||||||||||
селения |
популяции). |
|
|
|
|
|
|
|||||||||||||||||||
Часто оказывается так, что опре- |
|
|
|
|
|
|
||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||
деленный |
генотип |
|
имеет |
селективное |
|
Из других более частных форм |
||||||||||||||||||||
преимущество только тогда, когда он |
отбора |
|
упомянем |
дестабилизирующий |
||||||||||||||||||||||
редок в популяции, и немедленно те- |
отбор (Д. К. Беляев) и отбор место- |
|||||||||||||||||||||||||
ряет |
это |
преимущество, |
оказавшись |
обитания. |
Под |
дестабилизирующим |
||||||||||||||||||||
визбытке. Это типичная ситуация отбором имеется в виду разрушение
действия |
част от но зависимо го |
отбора. |
коррелятивных связей |
в организме |
||
Именно под таким отбором находятся |
при интенсивном отборе в каком-то |
|||||
многие признаки покровительственной |
определенном направлении. Так, на- |
|||||
окраски |
типа мимикрии (например, |
пример, отбор на уменьшение агрес- |
||||
при |
апостатическом |
отборе — одной |
сивности у хищных зверей в неволе |
|||
из |
форм |
частотнозависимого |
отбо- |
ведет к дестабилизации цикла размно- |
||
ра — полиморфизм в популяции жерт- |
жения (животные могут |
размножать- |
||||
вы может поддерживаться избиратель- |
ся круглый год). Последняя форма |
|||||
ным |
хищничеством, |
направленным на |
отбора может быть проиллюстрирова- |
|||
более многочисленную форму). |
|
на таким примером: самки двух близ- |
||||

i 22
i 22 Часть 3. Учение о м и к р о э в о л ю ц и и
Рис. 10.9. Пример полового диморфизма в строении приматов: Б—голова самца тамарина (Saguinus mystax); А—голова самца носача (Nasalis larvatus). Усы и нос в значительно меньшей степени развиты у самок этих видов (из А. Портмана, 1967; А. А. Парамонова, 1978)
ких видов дрозофил Гавайских остро- |
У самцов сильно развиты орудия для |
||||||||||||||||
вов откладывают яйца на одном и том |
турнирного |
боя |
(рога, |
|
клыки, |
шпоры |
|||||||||||
же виде растений, но самки одного |
и т. п.). Эти органы вначале возникли |
||||||||||||||||
вида откладывают яички на теневой |
как |
средство |
защиты |
в |
ходе |
борьбы |
|||||||||||
стороне растения, тогда как другого — |
за существование, а затем «подхва- |
||||||||||||||||
исключительно на |
солнечной стороне. |
тывались» половым отбором. В поло- |
|||||||||||||||
Отбор местообитаний определяет бо- |
вом отборе большую активность обыч- |
||||||||||||||||
лее полное использование пространст- |
но проявляют |
самцы, |
что |
выражается |
|||||||||||||
ва |
близкими |
симпатрическими |
видами |
в боях, пении, ухаживании, демонст- |
|||||||||||||
(ведет |
к |
увеличению |
суммы |
жизни, |
ративном поведении. Наиболее актив- |
||||||||||||
как образно заметил в свое время Ч. |
ные самцы чахце занимают типичные |
||||||||||||||||
Дарвин, не выделявший никаких спе- |
для вида территории и тем самым у |
||||||||||||||||
циальных |
форм естественного |
отбора, |
них больше шансов найти самку для |
||||||||||||||
но в своих работах прозорливо опи- |
спаривания. |
Развитие |
яркой |
окраски |
|||||||||||||
савший практически все основные фор- |
и привлечение внимания к самцам в |
||||||||||||||||
мы |
действия |
отбора). |
|
|
|
период брачного сезона для вида вы- |
|||||||||||
|
|
|
|
|
|
|
|
|
годно в целом, так как самки остаются |
||||||||
|
|
|
10.8. Половой |
отбор |
|
в относительной |
безопасности |
в труд- |
|||||||||
|
|
|
|
ный и ответственный период жизни, |
|||||||||||||
|
|
|
|
|
|
|
|
|
|||||||||
|
Естественный |
отбор, |
касающийся |
связанный |
с |
оставлением |
потомства. |
||||||||||
|
|
|
|
|
|
|
|
|
|
||||||||
признаков |
особей одного пола, назы- |
Половой |
|
отбор — не |
самостоя- |
||||||||||||
вается половым отбором. Обычно по- |
тельный фактор эволюции, а всего |
||||||||||||||||
ловой отбор вытекает из борьбы меж- |
лишь частный |
случай |
внутривидового |
||||||||||||||
ду |
самцами |
(а в |
более |
редких |
случа- |
естественного |
отбора. |
|
|
|
|
||||||
ях — между самками) за возможность |
|
|
|
|
|
|
|
|
|
||||||||
вступить |
в размножение. |
|
|
|
|
|
|
|
|
|
|
|
|||||
|
В теории естественного отбора Ч. |
10.9. |
Индивидуальный |
и |
групповой |
||||||||||||
Дарвин столкнулся с трудностью объ- |
|
|
|
|
©тбор |
|
|
|
|
||||||||
яснения |
фактов |
полового |
диморфиз- |
|
|
|
|
|
|
|
|
|
|||||
ма |
(рис. |
10.9). Казалось бы, яркая |
Как уже указывалось, естествен- |
||||||||||||||
окраска самцов многих птиц, зазыва- |
ный отбор подразделяется на индиви- |
||||||||||||||||
ющие брачные крики и специфические |
дуальный и групповой. Индивидуаль- |
||||||||||||||||
запахи самцов действуют демаскиру- |
ный отбор сводится к дифференциаль- |
||||||||||||||||
юще, но они имеют смысл для размно- |
ному |
размножению отдельных |
особей, |
||||||||||||||
жения, для успеха в половом отборе, |
обладающих преимуществами в борьбе |
||||||||||||||||
так как повышают возможность уча- |
за существование в пределах популя- |
||||||||||||||||
стия |
в |
воспроизведении |
потомства. |
ций. |
Индивидуальный |
отбор |
основан |
||||||||||
|
|
Глава |
10. |
Естественный |
отбор — д в и ж у щ а я и |
направляющая |
сила |
эволюции |
|
|
159 |
||||||||||||||
на соревновании особей внутри попу- |
давления отбора. Наблюдения показа- |
||||||||||||||||||||||||
ляций. |
|
|
|
|
|
|
|
|
|
|
|
ли, что виды антилоп африканских |
|||||||||||||
|
Естественный |
отбор, |
|
«перераба- |
саванн поедают разные части травя- |
||||||||||||||||||||
тывая» |
незначительные |
наследствен- |
нистых |
растений (одни |
едят |
только |
|||||||||||||||||||
ные |
различия |
особей |
и |
«складывая» |
мягкие верхушки трав с цветами, дру- |
||||||||||||||||||||
их в определенном направлении, спо- |
гие — только |
сухие |
соломинки |
трав, |
|||||||||||||||||||||
собствует |
постепенному |
отклонению |
третьи — колючие листья и т. п.). Та- |
||||||||||||||||||||||
потомков от предков. Любые признаки |
кое |
положение — результат |
|
группо- |
|||||||||||||||||||||
и свойства видов и более крупных так- |
вого отбора между близкими видами, |
||||||||||||||||||||||||
сонов формируются в процессе отбо- |
способствующий |
увеличению |
|
«суммы |
|||||||||||||||||||||
ра особей на основе оценки их инди- |
жизни» |
на |
единице |
площади. |
|
|
|
||||||||||||||||||
видуальных различий. На этом посто- |
|
Во всех без исключения случаях |
|||||||||||||||||||||||
янном |
фоне |
индивидуального |
|
отбора |
групповой отбор основан на внутри- |
||||||||||||||||||||
в |
природе |
и |
осуществляется |
|
группо- |
популяционном |
естественном |
|
отборе. |
||||||||||||||||
вой |
отбор |
— |
преимущественное |
раз- |
Это и понятно, так как конкуренция |
||||||||||||||||||||
множение |
особей |
какой-либо |
|
группы. |
видов в процессе эволюции осущест- |
||||||||||||||||||||
|
При групповом отборе в эволюции |
вляется через конкуренцию их инди- |
|||||||||||||||||||||||
закрепляются |
признаки, |
благоприят- |
видов. |
Возникновение эволюционных |
|||||||||||||||||||||
ные для группы, но не всегда благо- |
новшеств происходит только при ин- |
||||||||||||||||||||||||
приятные для особей. В групповом |
дивидуальном |
|
отборе |
бесчисленных |
|||||||||||||||||||||
отборе |
группы |
особей |
соревнуются |
вариантов, а групповой отбор выбира- |
|||||||||||||||||||||
друг с другом в создании и поддержа- |
ет уже из готовых приспособлений, |
||||||||||||||||||||||||
нии |
целостности |
надорганизменных |
возникших |
на |
внутривидовом |
уровне. |
|||||||||||||||||||
систем. |
|
|
|
|
|
|
|
|
|
|
|
Промежуточное |
положение |
занимает |
|||||||||||
|
Родственные особи обладают боль- |
групповой |
внутривидовой |
отбор — от- |
|||||||||||||||||||||
шим сходством генотипов, чем нерод- |
бор разных семей, популяций, групп |
||||||||||||||||||||||||
ственные. Поэтому если какой-либо |
популяций. |
|
|
|
|
|
|
|
|
||||||||||||||||
признак у части родственных особей |
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||
будет содействовать выживанию ближ- |
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||
них, |
то |
|
такой |
признак |
может |
закре- |
10.10. Творческая роль естественного |
||||||||||||||||||
питься |
в |
эволюции даже |
при |
условии |
|||||||||||||||||||||
|
|
|
|
отбора |
|
|
|
|
|||||||||||||||||
его отрицательного влияния на непо- |
|
|
|
|
|
|
|
|
|||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||
средственного |
носителя. |
|
Например, |
|
Оппоненты |
|
теории |
естественного |
|||||||||||||||||
особь, |
предупреждающая |
|
криком о |
отбора иногда говорят о тавтологич- |
|||||||||||||||||||||
появлении |
хищника |
своих |
родичей, |
ное™ |
этой |
концепции: |
«выживают |
||||||||||||||||||
чаще всего оказывается жертвой напа- |
наиболее |
приспособленные, |
так |
как |
|||||||||||||||||||||
дающего. Однако все остальные особи |
они наилучше приспособлены», т. е. |
||||||||||||||||||||||||
этой группы будут спасены, а посколь- |
отбор «выискивает» в популяциях уже |
||||||||||||||||||||||||
ку альтруистический признак характе- |
приспособленных и их-то и |
|
оставля- |
||||||||||||||||||||||
рен хотя бы и для некоторых из них |
ет для размножения. Такая постанов- |
||||||||||||||||||||||||
тоже, этот признак будет распростра- |
ка вопроса оказывается по существу |
||||||||||||||||||||||||
няться |
естественным |
отбором |
(отбо- |
антиисторической и искажающей ме- |
|||||||||||||||||||||
ром сородичей). Предполагается, что |
ханизм действия отбора. На самом де- |
||||||||||||||||||||||||
именно групповым отбором закреп- |
ле признаки (адаптации) никогда не |
||||||||||||||||||||||||
лены |
в |
эволюции |
свойства, |
связанные |
появляются в готовом виде. Естествен- |
||||||||||||||||||||
с |
регуляцией |
численности |
популяции. |
ный отбор ответственен за суммирова- |
|||||||||||||||||||||
Групповой отбор может привести к |
ние мелких отклонений. Антидарвинис- |
||||||||||||||||||||||||
вытеснению |
одной из |
конкурирующих |
ты приписывали отбору роль механи- |
||||||||||||||||||||||
групп и тем самым к уменьшению |
ческого сита. Согласно такому мнению |
||||||||||||||||||||||||
группового разнообразия, либо к воз- |
отбор не создает, а лишь «сортирует» |
||||||||||||||||||||||||
никновению |
новых |
различий |
|
между |
имеющиеся в |
популяциях изменения. |
|||||||||||||||||||
формами |
и |
тем самым |
к |
|
понижению |
Подобные |
суждения |
об |
отборе |
осно- |
|||||||||||||||
ваны на учете лишь элиминирующей |
двух букв легко могут быть дополнены |
|||||||||||||||||||||||
его |
функции. |
|
|
|
|
|
|
|
третьей недостающей буквой и приве- |
|||||||||||||||
|
Определение отбора как диффе- |
дут к получению слова «кот». Хотя мы |
||||||||||||||||||||||
ренциального |
размножения |
особей |
с |
и не создали букв, но добились их |
||||||||||||||||||||
разными генотипами тесно связано с |
смыслового сочетания. Точно так же и |
|||||||||||||||||||||||
признанием его творческой роли. Один |
признаки |
|
(приспособления) |
могут |
||||||||||||||||||||
и |
тот |
же |
материал |
(наследственная |
«складываться» |
под |
влиянием |
отбора |
||||||||||||||||
изменчивость) |
в зависимости |
от |
усло- |
в результате |
сочетания |
разных |
генов. |
|||||||||||||||||
вий и направления отбора может при- |
К этому следует добавить, что каждый |
|||||||||||||||||||||||
вести к различным адаптациям. В этом |
ген отбирается и накапливается не сам |
|||||||||||||||||||||||
отношении действие отбора по праву |
по себе, а в системе с другими гена- |
|||||||||||||||||||||||
сравнивают с деятельностью скульп- |
ми |
в |
целостном |
генотипе, |
как |
бук- |
||||||||||||||||||
тора. Как он из бесформенной глыбы |
вы |
в |
словах |
и |
|
слова |
в |
предложе- |
||||||||||||||||
гранита |
создает |
чудесные |
творения, |
ниях. |
|
|
|
|
|
|
|
|
|
|
|
|||||||||
«лишь» отсекая ненужное, так и отбор, |
Отбор формирует признаки и свой- |
|||||||||||||||||||||||
складывая |
бесчисленные |
изменения, |
ства |
путем |
интегрирования |
отдельных |
||||||||||||||||||
создает приспособления и виды, устра- |
случайных |
|
уклонений |
с |
незначитель- |
|||||||||||||||||||
няя от размножения «лишь» менее |
ной |
адаптивной |
ценностью. |
|
|
|
||||||||||||||||||
удачные особи. Аналогия со скульпто- |
Сходство и различие в действии |
|||||||||||||||||||||||
ром, конечно, неполная: у человека- |
естественного |
и |
искусственного |
отбо- |
||||||||||||||||||||
творца всегда есть план будущего тво- |
ра. Для пояснения творческой ролк |
|||||||||||||||||||||||
рения, |
и, |
отсекая |
те |
или |
иные |
куски |
естественного |
отбора |
можно сравнить |
|||||||||||||||
от глыбы гранита, он идет к осущест- |
его действие с действием искусствен- |
|||||||||||||||||||||||
влению цели. У отбора такой цели нет, |
ного отбора |
(как это делал еще Ч. Дар- |
||||||||||||||||||||||
направленность в действии отбора воз- |
вин) . |
|
|
|
|
|
|
|
|
|
|
|
||||||||||||
никает |
стихийно. |
Но |
у отбора |
есть |
Подобно |
|
тому |
как |
селекционер, |
|||||||||||||||
практически |
неограниченное |
время |
|
|||||||||||||||||||||
отбирая |
особей |
|
с |
незначительными |
||||||||||||||||||||
действия (миллионы и миллиарды лет) |
|
|||||||||||||||||||||||
полезными |
уклонениями |
в |
поколени- |
|||||||||||||||||||||
и |
огромное |
разнообразие |
материала |
|||||||||||||||||||||
ях, создает породы и сорта, так и есте- |
||||||||||||||||||||||||
для работы. |
|
|
|
|
|
|
|
|||||||||||||||||
|
|
|
|
|
|
|
ственный отбор ведет к образованию |
|||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||
|
Формирование |
любого |
|
генотипа |
видов в природе. Скорость работы |
|||||||||||||||||||
происходит под действием естествен- |
селекционера |
сравнительно высока, но |
||||||||||||||||||||||
ного отбора, что преобразует всю на- |
у естественного отбора есть то, чем |
|||||||||||||||||||||||
следственную |
изменчивость. |
Это, |
в |
селекционер |
|
не |
|
располагает,— прак- |
||||||||||||||||
свою очередь, ведет к изменению фе- |
тически неограниченное время дейст- |
|||||||||||||||||||||||
нотипа, т. е. к возникновению всех |
вия. |
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||
признаков и свойств организма. Пред- |
Несмотря на сходство в действии, |
|||||||||||||||||||||||
положим, что из большой выборки |
естественный |
и |
искусственный |
отбог |
||||||||||||||||||||
вырезанных букв алфавита, представ- |
различаются и по результатам. Искус- |
|||||||||||||||||||||||
ленных в равном числе, предстоит |
ственный отбор часто ведет к накопле- |
|||||||||||||||||||||||
сложить слово «кот». Для этого можно |
нию признаков, не приносящих польз; |
|||||||||||||||||||||||
извлечь буквы по 2 и 3 и практически |
для их обладателя |
(большинство видов |
||||||||||||||||||||||
никогда не получить нужного сочета- |
домашних |
|
животных |
и |
культурных |
|||||||||||||||||||
ния. Но если из множества вынутых |
растений не могут существовать поэ- |
|||||||||||||||||||||||
букв вернуть в ящик К, О и Т |
(со вре- |
тому без постоянной поддержки чело- |
||||||||||||||||||||||
менем в ящике останутся только эти |
веком в результате резко пониженной |
|||||||||||||||||||||||
буквы), то это повысит вероятность |
жизнеспособности в природных усло- |
|||||||||||||||||||||||
получения |
нужного |
сочетания |
букв. |
виях; рис. 10.10). Естественный отбор |
||||||||||||||||||||
Изменим еще одно условие: некоторые |
никогда не ведет к закреплению при- |
|||||||||||||||||||||||
из вынутых букв вернем в ящик попар- |
знаков, |
вредных |
для вида. |
|
|
|
||||||||||||||||||
но |
(КО, КТ, |
ОТ), а |
другие |
оставим |
Другое |
|
различие |
между |
искусст- |
|||||||||||||||
одиночными. |
Такие |
комбинации |
из |
|
||||||||||||||||||||
венным |
и |
естественным |
отбором |
за- |
||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||