

Вракин В.Ф, Сидорова М.В. |
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ |
|
|
и липидов образуют сложную структурную основу клетки; при уплотнении протоплазма способна образовывать новые белковые мицеллы и восстанавливать нарушение связи между ними, а при нарушении целостности клетки
— уплотняться на наружной поверхности с образованием мембранноповерхностной реакции преципитации. Поэтому клетка быстро ликвидирует свои повреждения.
В определенных условиях дисперсная фаза коллоида обособляется не в виде осадка, а в виде раствора. При этом коллоидный раствор расслаивается на два раствора разной концентрации. Это коацервация (coacervacio — нагромождение, собирание в кучу). Коацерваты имеют вид капель, веретен, слоев. В неоднородной по химическому составу протоплазме, представляющей собой сочетание различных, часто противоположно заряженных коллоидов, возникают сложные комплексные коацерваты.
МОРФОЛОГИЯ КЛЕТКИ
Клетка состоит из сложноорганизованного живого вещества— протоплазмы, разделенного с помощью мембран на цитоплазму и ядро. Ядро отграничено от цитоплазмы ядерной оболочкой и состоит из кариоплазмы. Клетка отделена от внешней среды клеточной оболочкой — плазматической мембраной (плазмо-или цитолеммой), которая в животных клетках очень тонкая и видна лишь в электронный микроскоп (рис. 1).
Размеры и форма клеток очень разнообразны. Самые маленькие клетки не превышают нескольких микрометров (малые лимфоциты, клетки-зерна мозжечка), самые большие достигают нескольких сантиметров (яйцеклетки птиц). По форме клетки бывают шаровидные, овальные, кубические, призматические, звездчатые, дисковидные, с разнообразными отростками и т. д. Форма клеток тесно связана с их функцией и с механическим воздействием окружающей среды. Клетки, обладающие амебовидной подвижностью (лейкоциты), способны менять свою форму. Размеры ядра и цитоплазмы обычно находятся в определенных соотношениях, характерных для той или иной ткани и отражающих состояние клетки. У молодых и активно функционирующих клеток ядра бывают обычно крупнее, чем у клеток того же типа, находящихся в покое или стареющих.
Электронная микроскопия показала, что ядро, цитоплазма и клеточная оболочка имеют сложное строение. Они выполняют разнообразные функции. Нормальная жизнедеятельность клетки возможна только при их сохранности и взаимодействии.
Клеточная оболочка — поверхностный аппарат клетки, в состав которого входят: плазматическая мембрана, надмембранный комплекс и субмембранный опорно-сократительный аппарат (рис. 2).
www.timacad.ru
Вракин В.Ф, Сидорова М.В. |
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ |
|
|
Рис. 1. Схема ультрамикроскопического строения животной клетки:
I — цитолемма: 1 — пиноцитозная вакуоль; 2 — плотный контакт; 3
— десмосома; 4 — контакт по типу замка; 5 —щелевой контакт; 6 — простой контакт; II — цитоплазма; 7 — микроворсинка; 8 —ресничка; 9 — центросома; 10 — микротрубочки; 11 — гранулярная эндоплазматическая сеть; 12 — агранулярная (гладкая) эндоплазматическая сеть; 13 — аппарат Гольджи; 14 — митохондрия; 15 — лизосома; 16 — фаголизосома; 17 — полирибосомы; 18 — секреторная вакуоль- 19 — включения гликогена; 20 — жировые включения; III — ядро; 21 — кариолемма; 22 — пора; 23 — кариоплазма; 24 — хроматин; 25 — ядрышко; а — вндоцитоз; б — экэоцитоз.
www.timacad.ru
Вракин В.Ф, Сидорова М.В. |
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ |
|
|
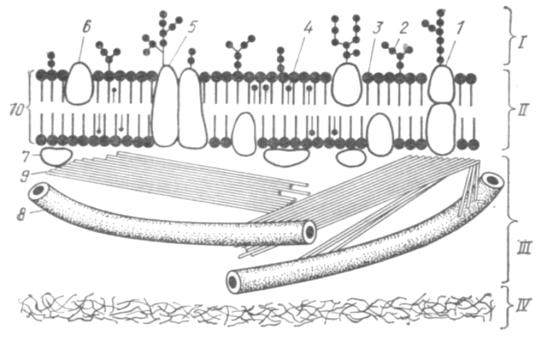
Рис. 2. Строение поверхностного аппарата клетки:
I — надмембранный комплекс (гликокаликс); II — плазмолемма; III — субмембранный комплекс; IV — цитоплазма; 1 — гликопротеиды; 2 — гликолипиды; 3 — фосфолипиды; 4 — холестерин; 5 — интегральные. 6 — полуинтегральные и 7 — периферические белки; 8 — микротрубочки; 9 — микрофиламенты; 10 — билипидный слой.
Плазматическая мембрана — плазмолемма или цитолемма имеет толщину около 10 нм и видна лишь в электронный микроскоп. Цитологические, биологические и физико-химические исследования показали, что она представляет собой белково-липидный комплекс, организованный определенным образом. Количество белков и липидов в плазмолемме по массе почти одинаковое, но мелких липидных молекул при этом оказывается гораздо больше, чем крупных белковых. Молекулы липидов полярны. У них незаряженные гидрофобные хвосты и заряженные гидрофильные головки. В цитолемме они располагаются в два слоя, гидрофобными хвостами друг к другу, а гидрофильными головками наружу. Такие же структуры образуют молекулы липидов и в неживых объектах, располагаясь на поверхности воды. Молекулы липидов, входящие в мембраны, разнообразны и очень подвижны. Вдоль мембраны они могут перемещаться со скоростью более миллиона раз в секунду, образуя как бы вечно подвижное липидное озеро, в котором плавают молекулы глобулярных белков.
Белки мембран являются ферментными, рецепторными и структурными. Они также могут перемещаться в мембране в процессе выполнения функции, в результате чего меняются свойства мембраны и ее функциональная активность. Молекулы белков располагаются в мембране как между молекулами липидов— интегральные белки, так и под ними — периферические белки. А так как молекулы белков гораздо крупнее липидных, интегральные белки выступают наружу, образуя поверхностные слои мембраны. Такая структура
www.timacad.ru
Вракин В.Ф, Сидорова М.В. |
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ |
|
|
плазмолеммы называется мозаичной. В электронном микроскопе она выглядит трехслойной. Два крайних электронно-плотных слоя соответствуют гидрофильным головкам липидных молекул и белковым глобулам, а средний электронно-прозрачный слой — гидрофобным концам липидных молекул. Мембраны такого строения принято называть элементарной биологической мембраной.
Надмембранный комплекс (гликокаликс) — в основном углеводной природы. Представлен он полисахаридами, образующими с белками мембраны сложные соединения — гликопротеиды, а с ее липидами — гликолипиды. Длинные ветвящиеся углеводные концы молекул гликопротеидов и гликолипидов соединены в вязкий желеобразный надмембранный чехол толщиной 3—4 нм, который служит своеобразной межклеточной смазкой, склеивающей клетки между собой. В нем снижается скорость диффузии различных веществ, определенным образом располагаются ферментные комплексы, принимающие участие во внеклеточном расщеплении высокополимерных веществ. Особенно хорошо развит полисахаридный слой в кишечном эпителии, где очень активны процессы пристеночного (внеклеточного) пищеварения и всасывания. Считается, что надмембранный комплекс выполняет рецепторную функцию, в том числе и тканевой совместимости, функцию «узнавания» клетками друг друга, так как благодаря разнообразию химических связей молекул углеводов «рисунок» поверхности клеток тканеспецифичен и даже индивидуален. Он может приводить в действие иммунные механизмы.
Субмембранный комплекс сосредоточен в кортикальном слое цитоплазмы, прилежащем в плазмолемме и тесно с ней связанном. Представляет специализированную периферическую часть опорно-сократительной системы клетки. Состоит из белков, собранных в нитевидные структуры разной величины: микрофиламенты (диаметром 5—7 нм), микрофибриллы (около 10 нм) и микротрубочки (диаметром 20—22 нм). Все они вступают друг с другом в сложные химические и структурные связи, образуя терминальную сеть. Среди белков терминальной сети основными являются актин, миозин, тубулин, динеин и некоторые другие. Они образуют системы, способные к скольжению, взаимному перемещению, в результате чего перемещаются структуры плазмолеммы, связанные с этими системами, то есть осуществляется движение клетки и ее частей. Субмембранный комплекс принимает участие в рецепции, трансмембранном транспорте, стабилизирует белки плазмолеммы и осуществляет их направленное (координированное) перемещение.
Функции плазмолеммы. Плазмолемма вместе с над- и субмембранным комплексами осуществляет многообразные функции: барьерную, транспортную, рецепторную, двигательную, межклеточных взаимодействий и др.
Барьерная функция, проникновение веществ в клетку. Плазмолемма,
одевая клетку, отграничивает ее от внешней среды, в результате чего вещества внутрь клетки проникают избирательно. С помощью диффузии проникает в клетку и из клетки лишь вода и некоторые растворенные в ней газы. По градиенту концентрации (пассивным транспортом) перемещаются некоторые ионы и мелкие органические молекулы, особенно жирорастворимые. Против
www.timacad.ru
Вракин В.Ф, Сидорова М.В. |
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ |
|
|
градиента концентрации (активным транспортом) поступает большинство ионов, мелких молекул неорганических и органических веществ (соли, сахара, аминокислоты и др.). Процессы активного транспорта происходят с затратой энергии. В нем принимают участие все элементы поверхностного аппарата клетки. Макромолекулы, их агрегаты и более крупные частицы поступают в клетку благодаря эндоцитозу. При этом частица сначала адсорбируется на мембране клетки, затем окружается ею и отшнуровывается от мембраны с образованием вакуоли. Содержимым вакуоли будет захваченная частица, а ее стенкой — участок плазмолеммы. Если поглощаются более мелкие частицы и растворы, говорят о пиноцитозе, если более крупные — о фагоцитозе. Процесс выведения из клетки крупных частиц, заключенных в вакуоль, называется экзоцитозом. При этом стенка вакуоли встраивается в плазмолемму и становится ее частью, а содержимое изливается наружу.
Рецепторную функцию клетка выполняет с помощью специальных белков плазмолеммы и элементов гликокаликса (надмембранного комплекса). Рецепторы клетки разнообразны и многочисленны, что позволяет клеткам осуществлять взаимные контакты, реагировать на биологически активные вещества, осуществлять иммунные реакции и т. д.
Движение плазмолеммы осуществляется с помощью субмембранного комплекса, в котором скапливаются микротрубочки и микрофиламенты. При передвижении лейкоцитов, в процессе пино-, фагоцитоза и других формах движения клетки происходит перераспределение опорно-сократимой системы субмембранного комплекса и осуществляется движение клетки и ее частей.
Межклеточные контакты — соединения клеток, особенно распространены в эпителиальных тканях, где клетки плотно прилежат друг к другу, но имеются и между клетками других тканей. В образовании межклеточных контактов принимают участие все слои клеточной оболочки (поверхностного аппарата клетки). Межклеточные контакты неоднородны по структуре и функции. В зависимости от особенностей строения и функционального назначения различают следующие виды контактов: 1) простой контакт— клетки прилежат друг к другу, но между ними сохраняется щель 15—20 нм. Такой контакт широко распространен в различных органах и тканях; 2) зубчатый контакт, или контакт по типу замка, — плазмолеммы клеток образуют выпячивания, входящие во впячивания соседних клеток. Этот тип соединения способствует креплению клеток в единый пласт и характерен для эпителиев; 3) десмосома, или зона слипания, — сложноорганизованный участок двух соседних клеток, предназначенный для плотного скрепления клеток. Десмосомы широко распространены в покровном эпителии, но имеются также в железистом, почечном, кишечном эпителиях, в гладкой и сердечной мышечных тканях; 4) плотный (замыкающий) контакт — зона плотного прилегания клеток, в которой происходит слияние их мембран. Плотный контакт обычно окаймляет апикальные полюсы клеток, изолирует внутреннюю среду организма от внешней и распространен в эпителиях, особенно в кишечном; 5) щелевой контакт — особый тип соединения клеток, способствующий прове-
www.timacad.ru

Вракин В.Ф, Сидорова М.В. |
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ |
|
|
дению ионов и низкомолекулярных соединений из одной клетки в другую, минуя межклеточные щели.
Цитоплазма
Цитоплазма — сложная динамическая, многокомпонентная система клетки, в которой происходят основные метаболические процессы. В цитоплазме различают гиалоплазму, органеллы и включения.
Рис. 3. Участок клетки с фибриллярными структурами цитоплазмы (схема):
1 — плазмолемма; 2— цистерна эндоплазматической сети; 3 — микротрубочка; 4— микрофиламенты; 5 — полисома; 6 — митохондрия; 7 — ее наружная и 8 — внутренняя мембраны; 9 — криста; 10 — матрикс; 11 — мицелла.
Гиалоплазма (hyaline — прозрачный)—жидкая внутренняя среда клетки, состоящая из воды, низкомолекулярных веществ, растворенных в воде, водорастворимых веществ и высокомолекулярных веществ, рассредоточенных в виде мицелл и нитей, образующих нежную трехмерную сеть, видимую в электронный микроскоп лишь при очень большом увеличении (рис. 3). Особенности клетки как коллоидной системы, ее осмотические и буферные свойства в основном определяются составом гиалоплазмы и процессами, происходящими в ней. Гиалоплазма содержит большое количество ферментов (до 25% белков клетки приходится на гиалоплазму), участвующих в процессах гликолиза, метаболизма сахаров, азотистых оснований, аминокислот, липидов. В гиалоплазме располагаются молекулы АТФ, транспортные РНК и ферменты активации аминокислот при синтезе белка. Через гиалоплазму осуществляется большая часть внутриклеточного транспорта ионов, аминокислот, жирных кислот, нуклеотидов, сахаров и других веществ. Здесь откладываются запасные питательные вещества.
www.timacad.ru
Вракин В.Ф, Сидорова М.В. |
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ |
|
|
Органеллы (маленькие органы) — постоянные составные части цитоплазмы, выполняющие определенные функции. Одни из органелл присутствуют в каждой клетке организма и поэтому называются общими, другие — лишь в клетках определенного типа и называются специальными. К общим органеллам относят митохондрии, цитоплазматнческую (эндоплазматическую) сеть, пластинчатый комплекс (аппарат Гольджи), лизосомы, рибосомы, центросому, микротрубочки, микрофибриллы, а к специальным — миофибриллы, нейрофибриллы, тонофибриллы, реснички, жгутики, микроворсинки.
Большинство общих органелл имеет мембранное строение, то есть представляет собой структуры, стенка которых образована элементарной биологической мембраной. Мембраны органелл устроены так же, как и плазмолемма, и выполняют те же функции: транспортную, разграничительную и интегрирующую.
Мембранные органеллы — это эндоплазматическая сеть, комплекс Гольджи, лизосомы и митохондрии.
Эндоплазматическая (цитоплазматическая) сеть — впервые описана в 1945 г. К- Портером с сотрудниками. Эта широко распространенная органелла состоит из вытянутых цистерн, трубочек, мешков, узких каналов, связанных взаимными переходами, в результате чего образуется единая внутриклеточная сеть, которая как бы делит цитоплазму на отсеки, участки. Но при этом не имеет непосредственных контактов с плазмолеммой. Данная органелла очень подвижная и ранимая, ее составные части легко отшнуровываются, фрагментируются, объединяются, но содержимое при этом не изливается в гиалоплазму, а остается окруженным мембранами цитоплазматической сети. Различают агранулярную (гладкую) и гранулярную (шероховатую) эндоплазматическую сеть (рис. 4). Цистерны гранулярной эндоплазмати-ческой сети имеют полости шириной от 20 нм до нескольких микрометров. Со стенками гранулярной эндоплазматической сети связаны рибосомы. Повидимому, первоначально в клетке появляется именно этот вид сети, и лишь в процессе дифференцировки некоторая часть цистерн теряет рибосомы и ветвится, превращаясь в гладкую эндоплазматическую сеть. Обратных переходов, образования из агранулярной эндоплазматической сети гранулярной ее разновидности не наблюдается. Элементы гладкой эндоплазматической сети уже, их диаметр до 100 нм. Они представляют собой сложную систему переплетающихся трубочек, канальцев и пузырьков небольшого диаметра.
Цитоплазматическая сеть — своеобразная транспортная система клетки, по которой перемещаются разнообразные вещества, однако механизм транспорта неясен. Большая поверхность мембран сети говорит о том, что в клетке может происходить одновременно большое количество разнообразных реакций.
Основная функция гранулярной цитоплазматической сети — синтез белков. На рибосомах, прикрепленных к ее цистернам, синтезируются белки секреторных гранул, ферментные белки лизосом и белки, входящие в состав мембран клетки. Все эти типы белков транспортируются по цистернам цитоплазматической сети, не выходя в гиалоплазму. В гранулярной цитоплазма-
www.timacad.ru

Вракин В.Ф, Сидорова М.В. |
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ |
|
|
тической сети синтезируются, кроме того, некоторые углеводные соединения и липиды, входящие в состав мембран. Синтезированные молекулы подвергаются различным воздействиям (переаминирование, фосфорилирование и др.), в результате чего меняются их свойства, образуются сложные комплексные соединения. Гранулярная сеть считается местом образования мембран для всей клетки. Войдет ли вновь созданная мембрана в состав стенки эндоплазматической сети, пластинчатого комплекса, лизосомы или плазмолеммы зависит от того, какие белки и фосфолипиды синтезированы гранулярной цитоплазматической сетью.
Рис. 4. Пластинчатый комплекс Гольджи и эндоплазматическая сеть:
1 — цистерны пластинчатого комплекса; 2— вакуоли; 3 — секреторные гранулы; 4 — цистерны гранулярной эндоплазматической сети; 5 — гладкая эндоплазматическая сеть; 6 — транспортный пузырек.
Гладкая цитоплазматическая сеть многофункциональна. Она является местом синтеза ряда углеводов и липидов; поглощает, накапливает и транспортирует ионы (например, ионы кальция в волокнах поперечнополосатой мышечной ткани); осуществляет детоксикацию вредных продуктов обмена благодаря определенному набору ферментов, входящих в состав ее мембран
www.timacad.ru
Вракин В.Ф, Сидорова М.В. |
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ |
|
|
(в печени). Этот вид цитоплазматической сети очень распространен в клетках надпочечников, семенников — органах, где образуются стероидные гормоны.
Комплекс Гольджи (аппарат Гольджи), пластинчатый комплекс был описан К. Гольджи в 1898 г. в виде рыхлой сети, элементы которой имеют вид палочек, запятых, крючочков. Так, эта органелла выглядит в световой микроскоп.
Электронная микроскопия показала, что в данный комплекс входят цистерны, вакуоли и пузырьки. Как правило, он представляет собой стопку из 5—10 плоских мешковидных цистерн, узких в центре (7—10 нм) и расширяющихся по краям. По периферии они могут переходить в сеть канальцев и заканчиваться вакуолями, которые отшнуровываются от пластинчатого комплекса. В клетке насчитывают от одного до нескольких десятков комплексов. Расположен комплекс обычно вокруг ядра или над ним. Все его структуры построены из элементарной биологической мембраны, но толщина их и химический состав различны. Наиболее тонкие стенки имеют цистерны, обращенные к ядру и цитоплазматической сети, и по химическому составу подобны последней. Наиболее толстые стенки у цистерн, обращенных к наружным частям клетки. Они по структурно-химической организации подобны цитолемме.
Пластинчатый комплекс связан со всеми структурами клетки: с поверхностным и ядерным аппаратами, с цитоплазматической сетью, лизосомами. Выяснено, что в нем накапливаются, конденсируются и созревают продукты, синтезированные в цитоплазматической сети. При этом происходит их дальнейшая химическая перестройка с образованием секреторных гранул. В цистернах пластинчатого комплекса синтезируются полисахариды и в соединении с белками образуют гликозаминогликаны, мукопротеиды, протеогликаны, иммуноглобулины и другие комплексные соединения. Здесь синтезируются составные части надмембранного комплекса, которые потом без изменения входят в его состав. С помощью отшнуровывающихся вакуолей выводятся готовые секреты за пределы клетки. Сами же вакуоли встраиваются в цитолемму и становятся ее частью. Пластинчатый комплекс — место образования лизосом — вакуолей, наполненных гидролитическими ферментами, и пероксисом — пузырьков, содержащих окислительные ферменты.
Лизосомы открыты в 1949 г. де Дювом. Это разнообразные вакуоли диаметром 0,1—0,4 мкм. Стенка у них образована элементарной биологической мембраной, а содержимое представляет собой скопление гидролитических ферментов. В лизосомах обнаружено около 60 различных гидролаз, действующих в кислой среде. Ферменты лизосом способны разрушать практически1 все природные органические полимеры (белки, углеводы и нуклеиновые кислоты), но не разрушают клетку и стенки лизосом потому, что находятся в неактивном состоянии. Лизосомы в клетке очень разнообразны по размерам и внутреннему содержимому. Связано это с изменением лизосом в процессе функционирования, а также, возможно, со специализацией лизосом, так как. они отличаются по набору гидролитических ферментов. Ферменты лизосом в
www.timacad.ru
Вракин В.Ф, Сидорова М.В. |
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ |
|
|
течение всей жизни клеток отграничены мембраной. Выполняя гидролитическую функцию, лизосома приближается к пиноцитозной или фагоцитозной вакуоли и сливается с ней. В результате образуется единая вакуоль, содержащая ферменты и фагоцитированный материал — вторичная лизосома, или
фагосома.
Лизосомы переваривают не только продукты пиноцитозных и фагоцитозных вакуолей, но и разрушенные органеллы клетки (фрагменты митохондрий, цитоплазматической сети и других мембран). Непереваренные остатки могут накапливаться в лизосомах в виде слоистых телец, пигментных скоплений или удаляются из клетки в процессе экзоцитоза. Лизосомы в клетке могут передвигаться. В данном процессе какое-то участие принимают микротрубочки, однако механизм перемещения лизосом неясен. Большое количество лизосом бывает в клетках, выполняющих секреторную, экскреторную, всасывательную и фагоцитарную функции.
Митохондрии (mitos — нить, chondrion — гранула, зерно) впервые описаны в 1850 г. Келликером под названием саркосом, так как он увидел их в мышечных волокнах. В 1880 г. Альтман вновь привлек к ним внимание, разработав метод специфической окраски, а в 1898 г. Бенда назвал их митохондриями.
Митохондрии видны в световой микроскоп в виде нитчатых, палочковидных, гантелевидных или зернистых образований диаметром 0,2—2 мкм и длиной 1—10 мкм. В пределах одной клетки они могут иметь разную форму. Количество их в клетках сильно варьирует и исчисляется от единиц до тысяч. В соматических клетках млекопитающих обычно насчитывают 500—1000 митохондрий, которые занимают более 20% объема цитоплазмы. В связи с развитием электронно-микроскопических исследований стали поступать сведения о том, что эти органеллы могут иметь, очень сложную ветвистую структуру, в результате чего одна митохондрия попадает в срез несколько раз и создает впечатление большего числа митохондрий, чем на самом деле. Так или иначе общая поверхность их огромна. В клетках печени, например, она в 4—5 раз превышает площадь поверхности плазматической мембраны. Располагаются митохондрии в клетках в различных местах. Они скапливаются в тех участках, где расходуется энергия
{под апикальным полюсом всасывающих клеток, у базального полюса клеток почечных канальцев и т. д.) или где находится энергетический субстрат (около липидных включений).
Независимо от формы и размеров общее строение у митохондрий одинаково в любых клетках. Они ограничены двумя мембранами толщиной по 7 нм. Между мембранами имеется межмембранное пространство шириной
10—20 нм. Полость митохондрии заполнена матриксом тонкозернистого строения. Мембраны различаются своими структурно-биохимическими и физическими свойствами. Наружная мембрана гладкая, не связана ни с какими другими мембранами клетки, имеет вид замкнутого мешка. Для нее характерна неспецифическая проницаемость, малое содержание белков и большое количество липидов. Внутренняя мембрана отличается высокоспецифичной
www.timacad.ru
Вракин В.Ф, Сидорова М.В. |
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ |
|
|
проницаемостью и высоким содержанием белков (до 75% вещества мембраны). У нее многочисленные выросты пластинчатой или трубчатой формы, направленные внутрь митохондрии — гребни, или кристы. Располагаются они как вдоль, так и поперек митохондрии, поэтому образуется подобие сети. Количество и величина крист заметно меняются в зависимости от функционального состояния и возраста митохондрии. У молодых и малоактивных митохондрий крист мало, и они короткие. У зрелых и функционально активных количество и размеры крист увеличиваются. На поверхности кристы, обращенной к матриксу, обнаружены грибовидные тельца. Матрикс митохондрии представляет собой коллоидную систему, содержащую большое количество макромолекул различной природы.
Функция митохондрии — синтез АТФ. Она определяется набором окислительным ферментов, находящихся во внутренней мембране и матриксе. В последнем сосредоточены ферменты цикла Кребса (трикарбоновых кислот) и окисления жирных кислот. Во внутренней мембране расположена цепь переноса электронов (дыхательная цепь) и многочисленные транспортные системы, а на грибовидных тельцах локализованы ферменты фосфорилирования (превращения АДФ в АТФ). Все эти ферментные системы могут синтезировать и накапливать макроэргические (высокоэнергоемкие) соединения (главным образом в виде АТФ), только образуя сложные молекулярные комплексы в составе внутренней мембраны или будучи тесно с ней связанными. В местах потребления энергии АТФ легко распадается также при участии ферментов митохондрий. При этом образуется свободная энергия для выполнения механической, химической, электрохимической и осмотической работ. Поэтому митохондрии образно называют «силовыми станциями» клетки. Помимо этого, они могут участвовать в транспорте воды, накоплении Са, Mg
иР, в синтезе стероидных веществ.
Вматриксе митохондрий выявляются тонкие нити и мелкие гранулы. Оказалось что нити — это собственная ДНК митохондрий, отличающаяся по составу и структуре от ДНК ядра, а мелкие гранулы — митохондриальные рибосомы. Митохондрии, таким образом, имеют свой автономный аппарат для синтеза собственных митохондриальных белков.
Изучение живых митохондрий с помощью фазово-контрастной микроскопии показало, что они очень динамичны: могут расти в длину, сжиматься, ветвиться, делиться — и все это за период меньше одной минуты. Жизнь митохондрии продолжается около 20 дней, после чего она утилизируется лизосомами. Гибнущие митохондрии сменяются новыми. Количество митохондрий возобновляется за счет роста и деления предшествующих митохондрий перетяжкой или почкованием.
Немембранные органеллы — это рибосомы, центросома, микротрубочки, микрофибриллы.
Рибосомы как маленькие плотные тельца были впервые описаны Паладе в 1955 г., а в 1958 г. Роберте дал им название «рибосома». Их можно увидеть только в электронный микроскоп.
www.timacad.ru
Вракин В.Ф, Сидорова М.В. |
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ |
|
|
Это округлые образования диаметром 10—30 нм, состоящие из двух субъединиц. Обе субъединицы образованы РНК и белками примерно в равных количествах, которые связаны в виде рибонуклеопротеидов (РНП). Субъединицы неравны по величине. Большая субъединица напоминает полусферу с тремя торчащими выступами, а меньшая — короткую изогнутую палочку с выступом и несколькими полостями.
Рибосома принимает участие в синтезе белка. На ней происходит сборка полипептидов и белков. При этом к малой субъединице присоединяется молекула иРНК, а к большой — молекулы тРНК Возможно такое присоединение молекул диктуется формой субъединиц (В. Я. Стельмашук). В то время когда не происходит синтеза белка, субъединицы, как правило, разобщены. Их объединение происходит под действием ионов магния. Рибосомы синтезируются в ядрышке, но функционируют в основном в цитоплазме. Здесь они располагаются в гиалоплазме — свободные рибосомы или прикрепляются к цистернам цитоплазматической сети—связанные рибосомы. Белки, синтезированные свободными и связанными рибосомами, имеют разную судьбу. Связанные рибосомы синтезируют белки, которые, как правило, попадают в цистерны цитоплазматической сети, оттуда — в пластинчатый комплекс и в составе вакуолей покидают клетку (секреторные белки) или становятся белками лизосом. Свободные рибосомы в процессе синтеза белка объединяются
вгруппы — полисомы (полирибосомы). Они синтезируют белки, входящие в состав гиалоплазмы, структурные белки немембранных органелл, субмембранного комплекса и другие пластические и ферментативные белки клетки.
Микротрубочки существуют во всех клетках, кроме бактерий и некоторых водорослей, как самостоятельные структуры или в составе немембранных органелл (кроме рибосом). Представляют собой длинные, тонкие, неветвящиеся полые цилиндры с диаметром 22—26 нм и стенкой, построенной из глобулярного белка тубулина. Микротрубочки очень лабильны. Под влиянием изменения внутриклеточной среды происходит постоянная их самосборка и саморазборка. В результате одновременного протекания этих процессов в разных участках цитоплазмы система микротрубочек в целом поддерживается в состоянии динамического равновесия и в интерфазной клетке имеет вид трехмерной сети. В делящейся клетке сеть микротрубочек цитоплазмы исчезает и вместо нее формируется веретено деления.
Внастоящее время микротрубочки относят к скелетным образованиям клетки, поскольку форма последней в значительной мере зависит от того, как
вней распределены эти органеллы. -С ними связывают транспортную функцию клеточной оболочки на том основании, что в большом количестве они обнаруживаются в субмембранном комплексе. Микротрубочки — основные составные части органелл, осуществляющих движение клетки и ее частей: центросомы, ресничек, жгутиков. Однако механизм движения их неясен, так как сами микротрубочки не обладают способностью к сокращению, а движение тем не менее осуществляется. Возможно, своим расположением они создают систему упорядоченного движения внутриклеточных компонентов, являются для них как бы рельсами.
www.timacad.ru

Вракин В.Ф, Сидорова М.В. МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Центросома (клеточный центр) — органелла, присущая животным клеткам. В клетках растений ее нет. Обнаружена и описана практически одновременно Флеммингом в 1875 г. и Бенсденом в 1876 г.
Центросома состоит из центриолей и центросферы (рис. 5). В состав центросомы обычно входят две центриоли (диплосома), перпендикулярно расположенные друг к другу. Каждая из них состоит из девяти триплетов (троек) микротрубочек, уложенных с помощью специальных образований в виде цилиндра шириной 0,15 мкм и длиной 0,3—0,5 мкм. Микротрубочки центриолей такого же строения, как и микротрубочки цитоплазмы, и отличаются от них лишь большой стабильностью. Одна из центриолей материнская, другая — дочерняя. На материнской центриоли имеется ряд до-
полнительных структур: сателлиты, при-
|
датки и др., которых нет на дочерней |
||||
|
центриоли. Центросфера — зона цито- |
||||
|
плазмы |
с |
расходящимися |
нитчатыми |
|
|
структурами в виде лучей, окружающая |
||||
|
центриоли. |
Электронная микроскопия |
|||
|
показала, что лучистое ее сияние — это |
||||
|
многочисленные микротрубочки, от- |
||||
|
ходящие от сателлитов и радиально рас- |
||||
|
ходящиеся вокруг центриолей. В реснит- |
||||
|
чатых |
клетках |
центриолей гораздо |
||
|
больше, так как в основании каждой рес- |
||||
|
нички расположена центриоль в виде ба- |
||||
Рис. 5. Центросома: |
зального тельца. |
|
|
||
1 — материнская центриоль; 2 |
Центриоли |
являются |
центрами |
||
— дочерняя центриоль; 3 — са- |
сборки микротрубочек цитоплазмы, мик- |
||||
теллит; 4 — микротрубочки, |
ротрубочек веретена деления, выполня- |
||||
образующие центросферу; 5 — |
ют роль базальных телец, от которых от- |
||||
триплет микротрубочек цен- |
растают реснички и жгутики. Тем не ме- |
||||
триоли. |
нее центросома до сих пор — самая за- |
||||
|
гадочная органелла клетки. |
Высказыва- |
|||
ется предположение, что центриоли выполняют функцию внутриклеточной рецепции (Ю. С. Ченцов). Тесная связь ее с ядром с помощью микрофиламентов до сих пор не получила никакого объяснения.
Микрофибриллы и микрофиламенты — нитчатые структуры различной длины, толщиной от 5 до 10 нм, образованные белками. Располагаются по всей цитоплазме, но особенно развиты у апикального полюса клетки, где образуют терминальную сеть. Выполняют опорную и двигательную функцию, в различных тканях образуя специальные органеллы.
Специальные органеллы — постоянные структуры, присущие лишь клеткам определенных тканей. К ним относятся реснички, жгутики, тонофибриллы, миофибриллы, нейрофибриллы, микроворсинки.
Реснички и жгутики — органеллы движения. Реснички развиты в клетках эпителия дыхательных путей и некоторых отделов половых трактов, а
www.timacad.ru
Вракин В.Ф, Сидорова М.В. |
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ |
|
|
жгутики имеются у спермиев. Структура ресничек и жгутиков сходна, отличаются они лишь размерами. Реснички имеют длину 5—10 мкм, жгутики— 150 мкм. И те и другие представляют собой длинные тонкие выросты цитоплазмы, покрытые цитолеммой, имеющие в основании базальное тельце, а по всей длине — аксонему. Базальное тельце представляет собой центриоль, у которой можно наблюдать дополнительные структуры, характерные для материнской центриоли, а иногда можно видеть и дочернюю центриоль, расположенную под прямым углом к материнской. Основная нить ресничек и жгутиков — аксонема организована подобно центриоли, но содержит девять дублетов (пар) периферических и две центральные микротрубочки. Периферические и центральные микротрубочки связаны между собой в единую подвижную систему. При движении ресничек и жгутиков происходит скольжение микротрубочек друг по другу, в результате чего органеллы изгибаются.
Тонофибриллы, миофибриллы, нейрофибриллы — разновидности микрофибрилл и микрофиламентов, характерные для клеток той или иной ткани. Тонофибриллы развиты в эпителиальных тканях, где образуют скелет клеток. Миофибриллы развиты в мышечных тканях, и определяют сократимость мышечных клеток и волокон, а нейрофибриллы — в нервных клетках. Предполагается, что они участвуют в токе аксоплазмы и проведении нервного импульса.
Микроворсинки — выросты цитоплазмы, одетые цитолеммой и содержащие внутри пучок микрофиламентов. Они увеличивают всасывательную поверхность клетки. Особенно хорошо развиты в кишечном эпителии.
Включения. Это необязательные компоненты клетки, появляющиеся и исчезающие в зависимости от интенсивности и характера обмена веществ в клетке и от условий существования организма. Включения имеют вид зерен, глыбок, капель, вакуолей, гранул различной величины и формы. Их химическая природа очень разнообразна. В зависимости от функционального назначения включения объединяют в группы: трофические включения, секреты и инкреты, пигменты, экскреты и др. Среди трофических включений (запасных питательных веществ) важную роль играют жиры и углеводы. Белки как трофические включения используются лишь в редких случаях (в яйцеклетках в виде желточных зерен). Пигментные включения придают клеткам и тканям определенную окраску. Секреты и инкреты, накапливаются в железистых клетках, так как являются специфическими продуктами их функциональной активности. Экскреты — конечные продукты жизнедеятельности клетки, подлежащие удалению из нее.
Ядро
Ядро — такая же необходимая составная часть клетки, как и цитоплазма. Цитоплазма без ядра может существовать какое-то время, но вскоре гибнет, а ядро без цитоплазмы погибает еще быстрее. Как оформленная составная часть клетки, ядро возникло не сразу. Существуют клетки, у которых ядерный материал не отделен от остальной протоплазмы. Это бактерии, не-
www.timacad.ru
Вракин В.Ф, Сидорова М.В. |
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ |
|
|
которые водоросли. Клетки без оформленного ядра называют прока-
риотическими, с оформленным ядром — эукариотическими.
Для большинства эукариотических клеток характерно наличие одного ядра, но бывают и многоядерные клетки (например, мегакарноциты — специализированные клетки костного мозга, остеокласты в костной ткани и др.). Многоядерными (обычно двуядерными) могут становиться клетки печени, надпочечников, обычно одноядерные. При этом резко возрастает функциональная активность клетки, заметно увеличиваются ее размеры.
Форма ядер обычно зависит от формы клетки. У плоских клеток уплощенные, у сильно вытянутых веретеновидные ядра, у равносторонних клеток они, как правило, округлые, а у цилиндрических— овальные. Встречаются ядра сегментированные (у зернистых лейкоцитов крови) и даже ветвистые (у шелкопряда).
В клетках существуют определенные соотношения между размерами ядра и цитоплазмы, характерные для видовой принадлежности организма, ткани, органа. Эти соотношения непостоянные, они могут варьировать в широких пределах в зависимости от возраста и функциональной активности организма, органа, клетки. В молодых малодифференцированных клетках ядра относительно крупнее, чем в зрелых специализированных. У активно функционирующих клеток по сравнению с недеятельными клетками того же типа ядра более крупные. Изменения размеров ядра и цитоплазмы диктуются особенностями внутриклеточного метаболизма. Высказывается предположение (Нейфах), что определенное соотношение размеров ядра и цитоплазмы, называемое критическим, является толчком к делению клетки.
Химический состав ядра заметно отличается от состава цитоплазмы. В нем содержится почти вся ДНК клетки, много основных белков типа гистонов и негистоновых глобулярных белков. РНК и липидов меньше, чем в цитоплазме. Из неорганических веществ большую роль играют вода, ионы Са и
Mg.
Функции ядра определяются наличием в нем ДНК, в которой содержится вся генетическая информация. В ядре происходит редупликация (удвоение) молекул ДНК, благодаря чему генетическая информация в полном объеме передается следующим генерациям клеток. В ДНК в виде последовательности нуклеотидов закодирована информация о первичной структуре белков, которая реализуется в процессе синтеза. В ядре синтезируются все три вида РНК: рРНК, иРНК, тРНК и происходит образование субъединиц рибосом. Следовательно, ядро создает весь аппарат синтеза белка и регулирует этот синтез как с качественной, так и с количественной стороны. А это определяет жизнедеятельность клетки, так как практически все функции в ней обеспечиваются белками. Это хорошо видно в опытах по пересадке ядер из клетки одного типа в клетку другого типа. Пересаженное ядро направляет развитие и характер метаболизма в клеткеСтроениев сторонуядрасвоего. Ядротипасостоит. из ядерной оболочки (кариолеммы), отделяющей его от цитоплазмы, хроматина, ядрышка и кариоплазмы (рис. 6).
www.timacad.ru

Вракин В.Ф, Сидорова М.В. МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Кариолемма (ядерная оболочка, кариотека) состоит из двух элементарных биологических мембран толщиной по 7 нм, разделенных промежутками 10—20 нм. Наружная мембрана по структурно-химической организации подобна мембранам гранулярной ци-
|
топлазматической сети. К ней при- |
|||
|
соединяются рибосомы, она может |
|||
|
образовывать выпячивания, отшну- |
|||
|
ровываться в цитоплазму. Внутрен- |
|||
|
няя мембрана гладкая, без выпячи- |
|||
|
ваний. К ней прикрепляются нити |
|||
|
хроматина. |
|
|
|
|
Особенность |
кариолеммы — |
||
|
наличие пор. Это округлые участки |
|||
|
диаметром 50—100 нм, где слива- |
|||
|
ются наружная и внутренняя мем- |
|||
|
браны. Они заполнены, как проб- |
|||
|
кой, фибриллярными и гло- |
|||
Рис. 6. Строение ядра: |
булярными белками, |
образующими |
||
1 — кариолемма; 2— пора с поровым |
поровый комплекс. Размеры пор |
|||
комплексом; 3— кариоплазма; 4— |
обычно постоянны, а их количество |
|||
конденсированный и 5 — диффузный |
может |
уменьшаться |
в процессе |
|
хроматин; 6 — ядрышко; 7 — рибо- |
дифференцировки и функциониро- |
|||
сомы; 8 — цистерны эндоплазма- |
вания клетки. В среднем приходит- |
|||
тической сети. |
ся по 10 пар на 1 мкм2 поверхности |
|||
|
ядра. |
Через поры |
осуществляется |
|
транспорт веществ между ядром и цитоплазмой, но, несмотря на довольно большие размеры, далеко не все молекулы могут пройти через них — поровый комплекс обладает избирательной проницаемостью. Он пропускает, например, такие крупные структуры, как белки-гистоны, субъединицы рибосом, но задерживает многие мелкие неорганические молекулы.
Хроматин не виден в живом ядре, но становится заметен в фиксированной интерфазной клетке в виде глыбок, нитей, скоплений, окрашивающихся основными красителями. Назван так потому, что хорошо воспринимает красители (греч. chroma — цвет).
Химический анализ показал, что хроматин представляет собой комплексное соединение ДНК и белков — дезоксирибонуклеопротеиды (ДНП). Следовательно, хроматин, это не что иное, как хромосомы в деконденсированном состоянии. Одна молекула ДНК соответствует одной хромосоме. Длина молекул ДНК очень большая — от 0,5 мкм до 7 см. У человека длина молекул ДНК всех хромосом одной клетки равна 170 см. Чем более рыхло уложены хромосомы, тем активнее происходит на них транскрипция РНК и редупликация ДНК. Деконденсация хромосом неодинаково выражена на всем протяжении ДНП. Более конденсированные участки окрашены сильнее и заметны в виде глыбок хроматина (гетерохроматин). Чем заметнее хроматин в ядре, тем он более конденсирован и менее активен. Недеятельный хро-
www.timacad.ru
Вракин В.Ф, Сидорова М.В. |
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ |
|
|
матин полностью конденсирован, например, в зрелых эритроцитах птиц, где, как известно, синтетические процессы отсутствуют. Полностью конденсирован хроматин и в хромосомах во время митоза.
Наиболее активные синтетические процессы происходят в участках диффузного хроматина (эухроматин)—в полностью деконденсированных участках хромосом. Отсюда ясно, что по виду и расположению хроматина можно судить об активности синтетических процессов не только в ядре, но и в клетке. Чем диффузнее хроматин, тем выше синтез РНК и активнее в клетке идет синтез белка. Такой вид хроматина характерен для молодых, способных к делению клеток. В процессе дифференцировки клетки, по мере ее специализации, увеличивается количество конденсированного хроматина — происходит выключение участков молекулы ДНК из процесса транскрипции. Часто такой конденсированный хроматин накапливается в непосредственной близости к кариолемме. В последние годы появились данные, говорящие в пользу того, что конденсированный хроматин прочно связан с внутренней мембраной кариолеммы. В результате этого хромосомы в ядре интерфазной клетки, по-видимому, не разбросаны как попало, каждая из них занимает определенное пространство.
Ядрышко — наиболее плотный участок ядра округлой формы, диаметром 1—5 мкм. Окрашивается ядрышко основными красителями. Обычно в ядре 1—2 ядрышка, но может быть и много десятков, как, например, в ядрах ооцитов рыб. Общий объем ядрышек составляет около 1,5% ядерного объема. Находится ядрышко в непосредственной близости к определенному участку хромосомы, так называемому ядрышковому организатору. У свиньи, например, он обнаружен на 8-й и 10-й хромосомах, В области ядрышкового организатора ДНК расположены гены, на которых происходит синтез рибосомной РНК — основного вида РНК ядрышек. В области ядрышка рРНК синтезируется, накапливается и соединяется с белками — образуются рибонуклеопротеиды (РНП). Здесь же РНП объединяются в субъединицы рибосом. Ядрышко — неоднородная структура. Химически оно состоит на 60— 90% из белков, 2—8% РНК, 5—12% ДНК В нем различают тонкие фибриллы и гранулы, заключенные в аморфное вещество. Считают, что фибриллы — это участки молекул ДНК области ядрышкового организатора и РНП, а гранулы — субъединицы рибосом.
Расположение компонентов ядрышка может быть различным, как и его размеры. Это зависит от активности синтеза, накопления и выведения в цитоплазму рРНК При активном синтезе рРНК ядрышко крупное, его фибриллы лежат рыхло, гранул много. При затухании синтеза рРНК размеры ядрышка уменьшаются, фибриллы лежат плотно, гранул мало или нет совсем. В период деления клетки ядрышко исчезает, его материал рассеивается в содержимом ядра между хромосомами. Есть данные, что материал ядрышка входит в состав матрикса хромосом и вместе с ними передается дочерним клеткам. По окончании митоза, по мере деконденсации хромосом, происходит высвобождение компонентов матрикса и организация новых ядрышек.
www.timacad.ru
Вракин В.Ф, Сидорова М.В. |
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ |
|
|
Кариоплазма — жидкая фаза ядра, где располагаются все его структуры. В ней содержится большое количество белков, образующих так называемый ядерный матрикс. Функции этих белков во многом неясны. Предполагают, что они играют важную роль в поддержании общей структуры ядра и участвуют в регулировании синтеза нуклеиновых кислот. В состав кариоплазмы входят и мелкие молекулы органической и неорганической природы.
Вопросы для самоконтроля 1. Дайте характеристику химического состава и физико-химических свойств клетки. 2. Каково строение и функции клеточной оболочки, органелл, ядра?
Глава 3. ЖИЗНЕДЕЯТЕЛЬНОСТЬ КЛЕТКИ ПРОЦЕССЫ, ПРОИСХОДЯЩИЕ В КЛЕТКЕ
Все живое неразрывно связано с внешней средой, с которой оно обменивается веществом и энергией, реагирует на внешние воздействия. Не является исключением и клетка. В основе любых проявлений жизни клеток лежит обмен веществ.
Обмен веществ. Это определенный порядок превращения веществ, направленный на сохранение, самообновление клетки и выполнение ею своих функций. Интенсивность и направленность обмена веществ клетки в многоклеточном организме в основном определяются надклеточными регуляторными системами: нервной и эндокринной. Но существуют и сложные внутриклеточные системы регуляции процессов жизнедеятельности, что хорошо видно, например, в таких процессах, как редупликация ДНК, синтез белка. В процессе обмена веществ поступающие в клетку химические соединения претерпевают различные превращения (гидролиз, синтез, окисление, восстановление, переаминирование и др.), в результате чего они либо используются как промежуточные метаболиты, либо входят в состав структур клетки, либо в состав продуцируемых ею продуктов. Конечные продукты обмена удаляются из клетки.
Внутриклеточная регуляция обмена веществ производится особыми белками—ферментами, обладающими высокой специфичностью. Они катализируют (ускоряют), как правило, только одну какую-то реакцию. Локализация ферментов в определенном порядке на многочисленных мембранах клеточных органелл приводит к образованию целых ферментных систем, которые и направляют обмен веществ, ускоряя течение реакций лишь в определенном направлении. Регуляция деятельности ферментных систем осуществляется как извне (гормонами и другими биологически активными веществами), так и изнутри (продуктами, синтезированными в процессе реакции). В каждой ферментативной системе, как правило, существуют ключевые ферменты, связанные с продуктами их катализа отрицательной обратной связью. Отрицательная обратная связь — это такая связь, когда фермент своей деятельностью стимулирует образование какого-либо продукта реакции, а продукт реакции, накапливаясь в клетке, тормозит активность фермента. Обмен
www.timacad.ru

Вракин В.Ф, Сидорова М.В. |
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ |
|
|
веществ между клеткой и средой складывается из трех этапов: 1) поступление веществ в клетку; 2) преобразование веществ в процессе внутриклеточного обмена; 3) выведение из клетки продуктов метаболизма.
Поступление веществ в клетку и выведение из клетки метаболитов происходит через цитолемму и при непосредственном ее участии. Наличие внутриклеточных каналов, соединяющих внешнюю среду с цитоплазмой и с ядром, минуя цитолемму, в настоящее время отрицается. В зависимости от механизма проникновения вещества различают: пассивный перенос через мембрану клетки — по градиенту концентрации без затраты энергии, активный перенос — против градиента концентрации с затратой энергии. Например, нормально функционирующая клетка стремится к накоплению ионов калия и выведению ионов натрия. В результате в клетке калия оказывается больше, а натрия меньше, чем в межклеточной среде, но калий продолжает поступать в клетку, а натрий — выводиться из нее благодаря существованию так называемого натриево-калиевого насоса— определенного механизма переноса ионов с использованием АТФ и цАМФ.
Макромолекулы и более крупные частицы вещества поступают в клетку с помощью эндоцитоза — процесса проникновения в клетку веществ в виде вакуоли, стенки которой образованы отшнуровавшимся участком плазмолеммы (рис. 7). Разновидностями эндоцитоза являются пино-
цитоз (pynein — нить) и фагоцитоз
(phagein — пожирать). Пиноцитоз
— проникновение в клетку более мелких и жидких частиц, фагоцитоз
— более крупных и плотных частиц. Фагоцитоз у лейкоцитов был подробно изучен И. И. Мечниковым. Обычно при пиноцитозе возникает углубление цитолеммы, в которой оказывается захватываемый клеткой материал. Углубление затем принимает вид мешочка, потом пузырька, наконец, отшнуровывается от цитолеммы и погружается в цитоплазму клетки. Обра-
зуется пиноцитозная вакуоль. При фагоцитозе клетка для поглощения материала может образовывать как впячивания, так и выпячивания в виде ложноножек или парусов, которые, обволакивая, накрывают собой частоту вещества, погибшую
www.timacad.ru
Вракин В.Ф, Сидорова М.В. |
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ |
|
|
клетку, бактерию и, таким образом, погружают ее в цитоплазму. Образуется фагоцитарная вакуоль. Судьба пиноцитозной и фагоцитарной вакуолей сходна. И та и другая сливаются с лизосомами. Образуется фаголизосома, или вторичная лизосома, в которой происходит гидролиз (переваривание) захваченной частицы. Продукты гидролиза — мелкие молекулы простых веществ (сахара, аминокислоты, ионы, нуклеотиды и др.) — выходят через стенку этой пищеварительной вакуоли в гиалоплазму, а лизосома с оставшимися непереваренными продуктами образуют остаточное тельце, подлежащее удалению из клетки. Процесс выведения веществ из клетки называется экзоцитозом. Он подобен эндоцитозу, но идет в обратном направлении. Иногда при нарушении обмена веществ в стареющем организме можно видеть накопление в клетке остаточных телец в виде миелиновых телец, липофусциновых гранул, молочных камней и других образований, нарушающих жизнедеятельность клетки. Энергия, необходимая для осуществления процессов поглощения и переваривания веществ, поступает из митохондрий в виде АТФ.
Использование веществ, поступающих в процессе внутриклеточного обмена можно проследить на примере синтеза белков и образования секретов (см. «Железистые эпителии»).
Синтез белков (рис. 8) складывается из нескольких этапов: активации, элонгации и терминации. Первичная структура белка— последовательность аминокислот в белковой цепи детерминируется (определяется) структурой ДНК. Участок молекулы ДНК, ответственный за синтез одного вида белка, называется геном. В период активного белкового синтеза отдельные участки молекул ДНК деконденсируются, спирали ее расходятся и на них по принципу комплементарности под действием ферментов РНК — полимераз синтезируются молекулы иРНК из запаса нуклеотидов, сахаров, фосфатов, азотистых оснований, имеющихся в ядре. Этот процесс называется матричным синтезом или транскрипцией (переписывание). Ясно, что нуклеотидная последовательность иРНК точно такая же, как и участка ДНК, на котором она синтезирована. С одного участка ДНК может сниматься много одинаковых иРНК. Все они поступают в цитоплазму и вступают в тесную связь с меньшей субъединицей рибосомы. Это стимулирует сборку рибосом и полирибосом. Чем больше рибосом вошло в состав полирибосомы, тем больше синтезируется белковых молекул.
Наряду с этим в цитоплазме происходит активация аминокислот— соединение аминокислот с АТФ и присоединение активированных аминокислот к транспортным РНК. Каждой аминокислоте соответствует своя тРНК и свой фермент, активирующий эту реакцию. тРНК — небольшая молекула, на одном конце которой имеется антикодон (триплет)—участок из трех азотистых оснований, комплементарный определенному кодону иРНК. тРНК переносит соединенную с ней аминокислоту на рибосому и прикрепляется к большой субъединице. При этом антикодон тРНК на короткое время присоединяется к триплету иРНК по принципу комплементарности. иРНК делает один шаг на рибосоме, аминокислота оказывается присоединенной к строя-
www.timacad.ru

Вракин В.Ф, Сидорова М.В. |
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ |
|
|
щейся полипептидной цепочке, а тРНК, освобождаясь от связи с рибосомой, аминокислотой и иРНК, вновь попадает в гиалоплазму и может присоединять следующую молекулу аминокислоты. иРНК протягивается через рибосому, триплет за триплетом, в результате чего идет наращивание белковой молекулы, пока вся иРНК не будет прочитана.
Процесс прочтения информации, закодированной в иРНК, на рибосоме с помощью тРНК называется трансляцией. Синтезированная белковая молекула попадает либо в гиалоплазму, либо в цистерны цитоплазматической сети и приобретает специфическую форму.
Считается, что белки, попавшие в гиалоплазму, используются внутри клетки. Это могут быть структурные белки немембранных органелл, субмембранного комплекса, а также пластические и ферментные. Белки, поступающие в цистерны цитоплазматической сети, являются белками мембран клетки, лизосом и секреторных гранул, которые удаляются
из клетки.
Кругооборот мембран цитоплазмы. Многие процессы клетки, такие, как пиноцитоз, фагоцитоз, экзоцитоз, секреция, экскреция связаны с использованием мембранных структур в качестве упаковки того или иного содержимого. Можно было бы предположить большой расход плазмолеммы у всасывающих клеток или, наоборот, приращение ее у секретирующих. Однако большинство клеток в процессе жизнедеятельности мало меняет свои размеры. Объясняется это
тем, что процессы катаболизма и анаболизма в клетке тесно связаны между собой и с кругооборотом мембранных структур. Источником всех мембран клетки считается гранулярная цитоплазматическая сеть. В процессе транспорта веществ ее элементы отрываются и переходят в состав пластинчатого комплекса. Там они видоизменяются, приобретают сходство с клеточной мембраной, а их содержимое созревает, превращаясь в секреторную или экскреторную гранулу. Зрелая гранула в составе вакуоли отрывается от пластинчатого комплекса и, подходя к клеточной мембране, изливается за пре-
www.timacad.ru
Вракин В.Ф, Сидорова М.В. |
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ |
|
|
делы клетки. Оболочка вакуоли встраивается в цитолемму. Параллельно с этим участки цитолеммы обособляются в качестве оболочки пиноцитозной или фагоцитарной вакуоли, попадают в цитоплазму и подвергаются гидролизу с помощью лизосом. При этом разбирается и оболочка вакуоли (бывший участок цитолеммы). Из ее белков синтезируются мембраны цитоплазматической сети, которые затем в районе пластинчатого комплекса вновь видоизменяются. Такой кругооборот мембран существует в течение всего периода активного функционирования клетки.
Реакции клетки на изменение внешней среды. Клетки обладают раз-
дражимостью — специфической реакцией на воздействия внешней среды. Характер реакции тканеспецифичен. В ответ на одно и то же, например, механическое воздействие мышечная клетка отреагирует сокращением, нервная
— генерацией нервного импульса, мерцательный эпителий —движением ресничек. В организме есть клетки, реагирующие на определенные химические вещества активным движением, — лейкоциты.
В то же время на разнообразные воздействия физических и химических факторов (изменения химического состава поступающих в клетку веществ, изменение рН среды, колебания температуры, воздействие лучистой энергии, механические и др.) клетки разных тканей и органов реагируют однотипно. Реакция клеток зависит не от их тканевой принадлежности, а от силы и длительности действия фактора. Различают нормальную реакцию клетки, паранекроз (или некробиоз) и некроз.
Нормальная реакция клетки может не проявиться в ее морфологических изменениях, но может выразиться в некоторых изменениях размеров, структуры ядра и органелл, степени желатинизации цитоплазмы. При этом клетка переходит в более или менее активное состояние. При активации клетки (под действием гормонов, после кормления животного, при кратковременном стрессе, действии малых доз УФ-лучей и др.) в ней диспергируется хроматин, увеличивается ядрышко, растет число митохондрий и крист в них, лизосом, цистерн пластинчатого комплекса и цитоплазматической сети, усложняется структура митохондрий, уплотняется их матрикс. Снижение функциональной активности сопровождается конденсацией хроматина, уменьшением ядрышка и числа органелл. При этом митохондрии набухают, кристы исчезают, матрикс их становится светлее, в мембранах цистерн эндоплазматической сети появляются везикулы.
При более сильном и продолжительном воздействии раздражителя в клетке развивается состояние паранекроза, при котором значительно нарушаются функции, структура и химизм клетки. При паранекротическом состоянии уменьшается дисперсность коллоидов и возрастает вязкость ядра и цитоплазмы. Цитоплазма при этом мутнеет, а в ядре появляются нитчатые и глыбчатые структуры. Реакция цитоплазмы сдвигается в кислую сторону. Клетка теряет способность связывать прижизненные красители, и они диффузно окрашивают цитоплазму. Такие изменения ставят клетку на грань между жизнью и смертью. Но с прекращением действия раздражителя, если не повреждены структуры ядра, клетка возращается к нормальному морфофунк-
www.timacad.ru