

Вракин В.Ф, Сидорова М.В. |
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ |
|
|
клеток крови. 6. Чем лимфа отличается от крови? 7. Каково происхождение, строение, распространение в организме и функции рыхлой соединительной ткани? 8. Какие виды клеток и межклеточного вещества встречаются в соединительной ткани? Их строение и функции. 9. Какие виды жировой ткани вы знаете и чем отличаются друг от друга? 10. Виды хрящевой ткани, их строение, распространение и различия? 11. Развитие, строение и перестройка пластинчатой костной ткани. Где она находится?
Глава 9. МЫШЕЧНЫЕ ТКАНИ
К этому типу относятся ткани, основным свойством которых является способность к сокращению. Они обеспечивают движение животных в пространстве и двигательные функции органов (сердца, кишечника, мочеточников и др.). Мышечные ткани различают по происхождению, строению, особенностям функционирования. Общее у них — сходная форма эмбриональных клеток— миобластои и наличие большого количества специальных органелл — миофибрилл. В их состав входят распространенные в животных клетках сократительные белки (актин и миозин). Последние участвуют в движении клетки (лейкоциты) и ее частей (хвост спермия, отростки миоэпителиальных клеток, реснички эпителия), однако только в мышечных тканях эти белки образуют специфический актомиозиновый комплекс, наилучшим образом осуществляющий функцию движения. Мышечные ткани как более специализированные в филогенезе возникли позже эпителиальных и соединительных. Они способны к возбуждению, вызывающему их сокращение.
Кроме собственно мышечных структур, в состав мышечных тканей всегда входят нервные и соединительнотканные элементы. На основании особенностей строения и функции мышечные ткани позвоночных делят на гладкую и поперечнополосатые— скелетную и сердечную.
ГЛАДКАЯ (НЕИСЧЕРЧЕННАЯ) МЫШЕЧНАЯ ТКАНЬ
Гладкая мышечная ткань образует мышечную оболочку трубкообразных органов пищеварения (кроме глотки и части пищевода), дыхания, выделения, размножения, находится в стенках кровеносных сосудов, протоков желез, в селезенке, коже, ресничном теле глаза и в других органах.
Основная структурная единица гладкой мышечной ткани — клетка — гладкий миоцит. Это сильно вытянутая веретеновидная, иногда отростчатая (в мочевом пузыре, в эндокарде) клетка длиной 20—500 мкм (в матке беременных животных) и шириной 6—20 мкм. Вытянутое ядро находится в средней, утолщенной ее части. Здесь же, вокруг ядра, располагаются общие органеллы: митохондрии, комплекс Гольджи, центросома, рибосомы, эндоплазматическая сеть и включения гликогена. По всей клетке разбросаны протофибриллы — нитчатые структуры, образованные актином или миозином. Актиновые протофибриллы тоньше миозиновых и их в несколько раз (в 3—24 раза) больше. В расслабленной клетке протофибриллы ориентированы в ос-
www.timacad.ru
Вракин В.Ф, Сидорова М.В. |
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ |
|
|
новном продольно, но не образуют миофибриллярных комплексов. При сокращении образуется миофибриллярный актомиозиновый комплекс, приводящий к укорочению клетки.
Оболочка гладких миоцитов двухслойная. Она состоит из плазмолеммы обычного строения и базальной мембраны (пластинки) углеводно-белковой природы, выполняющей опорную функцию. Между ними имеется пространство шириной 15— 20 нм.
Функциональная единица гладкой мышечной ткани — пучок из 10—15 миоцитов, связанных с нервным волокном. В пучке мышечные клетки тесно связаны между собой с помощью десмосом, плотных и щелевых контактов. Это позволяет всем клеткам пучка одновременно реагировать на нервное раздражение, несмотря на то, что нервное окончание имеется лишь на одной из его клеток. Между мышечными клетками даже внутри пучка залегают тонкие коллагеновые и эластические волокна. Коллагеновые волокна оплетают миоциты, вплетаются в базальную пластинку, тем самым удерживая клетки от чрезмерного растяжения или сжатия. Пучки клеток отделены друг от друга прослойками соединительной ткани, в которой проходят сосуды и нервы.
Гладкая мышечная ткань иннервируется вегетативной нервной системой. Регуляция ее деятельности контролируется корой полушарий, но без участия сознания, сокращения осуществляются непроизвольно. Сокращается гладкая мышечная ткань медленно, ритмично. Период одного сокращения длится от 3 с до 5 мин. Может длительное время находиться в состоянии сокращения без заметного утомления. Такой характер сокращения называется
тоническим.
Происходит гладкая мышечная ткань из мезенхимы, клетки которой, дифференцируясь в гладкие миоциты, удлиняются и прилегают плотно друг к другу. Гладкие миоциты сохраняют способность к делению. Кроме того, на протяжении всего онтогенеза гладкие миоциты могут образовываться из недифференцированных клеток соединительной ткани, сопровождающих кровеносные сосуды.
ПОПЕРЕЧНОПОЛОСАТАЯ (ИСЧЕРЧЕННАЯ) СКЕЛЕТНАЯ МЫШЕЧНАЯ ТКАНЬ
Из данной ткани состоит скелетная мускулатура, мышцы языка, гортани, глотки, передней части пищевода, диафрагмы, глазного яблока, уха и некоторых других органов. Основная функция этой ткани — движение животного в пространстве и движение отдельных частей тела. Кроме того, она является основным источником тепла в организме, так как только 30—40% энергии мышечного сокращения используется в виде механической работы (движение), остальное превращается в тепловую энергию.
Структурная и функциональная единица поперечнополосатой скелетной мышечной ткани — мышечное волокно (рис. 31).
www.timacad.ru
Вракин В.Ф, Сидорова М.В. |
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ |
|
|
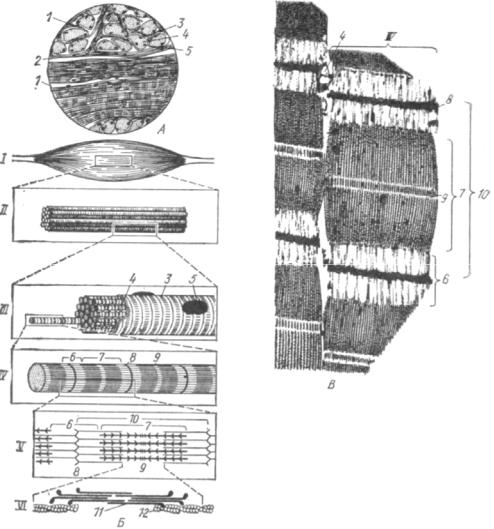
Рис. 31. Строение поперечнополосатой мышечной ткани:
А — гистосрез; Б — схема; В — электроннограмма участка мышечного волокна; I — мышца; II — пучок мышечных волокон; III — мышечное волокно; IV— миофибрилла; V — саркомер; VI — актомиозиновый комплекс:1 — эндомизий В; J — пернмизий; S — сарколемма; 4 — саркоплазма; 5 — ядро; 6 — И-диск; 7 — А-диск; 8 — Z-полоска (телофрагма); 9 — М-полоска (мезофрагма); 10 — саркомер; 11 — протофибрилла из молекул миозина; 12 — протофибрилла из молекул актина.
Это крупное симпластическое образование цилиндрической формы шириной 15—150 мкм (у крупного рогатого скота 16—95 мкм, у свиньи 20—90, у овцы 20—50, у лошади 50—80 мкм) и длиной от 1—2 мм до 10—15 см, с закругленными концами. Толщина волокон, возможно, ограничивается условиями их питания. Несмотря на то, что кровеносные капилляры прилегают к волокнам очень плотно, центральные участки слишком толстых волокон могут страдать от недостатка трофических и энергетических веществ. Оболочка волокна называется сарколеммой (sarcos — мясо), а содержимое — саркоплазмой. В волокне имеется от нескольких десятков до нескольких сотен ядер, которые у млекопитающих располагаются под сарколеммой. Ядра могут быть округлой, овальной и удлиненной формы.
www.timacad.ru
Вракин В.Ф, Сидорова М.В. |
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ |
|
|
Форма ядер зависит от возраста животного (у молодых животных ядра более округлые), степени сокращения волокна (в сокращенных волокнах ядра короче), от характера работы мышц. Если мышца с ярко выраженной динамической функцией, ее ядра более округлой конфигурации и лежат скоплениями. У мышцы со статической функцией ядра удлиненные, лежат цепочкой. В световом микроскопе каждое мышечное волокно выглядит исчерченным поперечными чередующимися светлыми и темными полосами, за что ткань получила свое название.
Мышечные волокна объединяются в пучки I порядка с помощью тонких прослоек рыхлой соединительной ткани (эндомизий). В эндомизии эластические волокна преобладают над коллагеновыми. В нем проходят кровеносные и лимфатические капилляры и нервные волокна. На одно мышечное волокно, как правило, приходится один капилляр, идущий вдоль волокна.
Пучки I порядка объединяются соединительной тканью в пучки II порядка, а те — в пучки III порядка. Эти прослойки соединительной ткани называют перимизием, а соединительную ткань, одевающую мышцу сверху, — эпимизием. В перимизии и эпимизии проходят крупные эластические и коллагеновые волокна, встречаются скопления жировых клеток. Мышечные волокна очень прочно связаны с одевающим их соединительнотканным каркасом. В местах соединения в мышечных волокнах образуются впячивания и складки, куда вклиниваются коллагеновые волокна и вплетаются там в сарколемму мышечных волокон.
Субмикроскопическое строение мышечного волокна. Сарколемма мышечного волокна состоит из плазмолеммы толщиной 7—10 нм и базальной мембраны (пластинки) толщиной 15— 50 нм. Плазмолемма через равные промежутки образует глубокие впячивания в виде трубок диаметром 4 нм (у свиньи)„ идущих поперек волокна и названных Т-каналами. Они играют большую роль в обмене веществ и проведения нервного импульса. Базальная мембрана (пластинка) выполняет опорную функцию и принимает участие в поддержании гомеостаза волокна. С ней связаны коллагеновые и эластические волокна, оплетающие вдоль и поперек мышечное волокно, образующие его внешний каркас.
В ядрах хроматин в виде глыбок. Во время сокращения ядра могут принимать штопорообразную форму, в них видны глубокие впячивания кариолеммы. В саркоплазме под сарколеммой вокруг ядер локализуются элементы гранулярной эндоплазматической сети, пластинчатого комплекса. В ней же находится пигмент миоглобин, который (как и гемоглобин в крови) связывает и депонирует кислород, поступающий в волокно, и отдает его при работе мышцы. Чем больше миоглобина в саркоплазме, тем больший резерв кислорода в мышце. Значительная часть саркоплазмы занята миофибриллами — специальными органеллами, ответственными за сокращение волокна. Вокруг них лежат митохондрии (саркосомы), гладкая эндоплазматическая (саркоплазматическая) сеть, включения гликогена.
Миофибрилла представляет собой нитчатую структуру с диаметром 0,5—2 мкм и длиной, соответствующей длине волокна, построенную из упо-
www.timacad.ru
Вракин В.Ф, Сидорова М.В. |
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ |
|
|
рядочение расположенных белков актина, миозина, тропомиозина, тропонина, актинина. По всей длине миофибриллы чередуются участки из белков актина и миозина. Миозин обладает двойным лучепреломлением (анизотропией), поэтому участки миофибриллы, построенные из этого белка, названы анизотропными дисками (А-диски). В световом микроскопе при проходящем свете они выглядят темными. Участки, построенные из актина, не обладают двойным лучепреломлением— это изотропные диски (И-диски). Они под микроскопом светлые. А-диски соседних миофибрилл волокна (так же, как и И-диски) расположены на одном уровне, в результате чего они воспринимаются как одна полоса, светлая или темная. От этого все волокно кажется по- перечно-исчерченным чередующимися темными и светлыми полосами.
Электронная микроскопия показала, что диски миофибрилл состоят из параллельно лежащих нитей, более толстых в А-диске и более тонких в И- диске. Нити эти названы протофибриллами. Тонкие протофибриллы построены из глобулярных молекул белка актина. Молекулы актина образуют цепочку, подобную двойной нитке бус, скрученной в виде веревочки, длиной 1 мкм и шириной 5—7 нм. Толстые протофибриллы построены ив молекул фибриллярного белка миозина, уложенных в виде пучка длиной 1,5—2 мкм и толщиной 10—25 нм.
Середину каждого И-диска пересекает темная Z-полоска, или телофрагма, а в средней части А-диска имеется светлая Н-зона, в центре которой проходит М-полоска, или мезофрагма. Z- и М-полоски имеют сложное, не до конца выясненное строение и выполняют опорную функцию, способствуя закреплению протофибрилл в таком положении, при котором каждая толстая про-тофибрилла оказывается окруженной шестью тонкими, а вокруг каждой тонкой лежат три толстые протофибриллы.
Тонкие протофибриллы одним своим концом прикреплены в области Z- полоски, другим — заходят в А-диск между толстыми протофибриллами, вплоть до Н-зоны. Следовательно, краевые участки А-диска образованы как миозиновыми, так и актиновыми протофибриллами, где они перекрывают друг друга, образуя зону перекрытия. Лишь в Н-зоне нет актиновых протофибрилл.
При мацерапии волокна или воздействием на него трипсином его миофибриллы распадаются по Z-полоскам на отдельные кусочки, названные саркомерами. Следовательно, саркомер — элементарная структурная единица миофибриллы. Его границами являются две соседние Z-полоски. Состоит он из половины И-диска, целого А-диска и половины другого И-диска, длина его 2,5—4 мкм. При сокращении происходит взаимное скольжение протофибрилл. Тонкие протофибриллы еще глубже втягиваются между толстыми, И-диск и Н-зона становятся уже. Длина саркомера уменьшается на 20—40%. При сильном сокращении И-диск и Н-зона исчезает совсем. Вся миофибрилла становится темной. Такую картину можно наблюдать в узлах сокращения
— местах наибольшего сокращения волокна.
Во время скольжения происходит перестройка (конформационные реакции) всех белков, входящих в состав миофибриллы, и между протофибрил-
www.timacad.ru
Вракин В.Ф, Сидорова М.В. |
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ |
|
|
лами возникают временные связи в виде мостиков, которые позволяют удерживать миофибриллы в их новом положении — образуется актомиозиновый комплекс. Процессы эти протекают со значительной затратой энергии и при обязательном присутствии ионов Са++ и Mg—Са — АТФазы.
По наиболее обоснованной теории сокращения, предложенной Р. Дэвисом, которая учитывает процессы превращения энергии, данные электронной микроскопии и рентгеноструктурного анализа, мышечные волокна сокращаются следующим образом. Нервное возбуждение через двигательное нервное окончание достигает сарколеммы. При этом происходит деполяризация мембраны волокна, которая с помощью Т-каналов быстро распространяется по всему волокну и приводит к изменению направления ионных потоков — возникает ток действия. Т-каналы тесно связаны с цистернами саркоплазматической сети. На уровне границы А- и И-дисков (у насекомых, рептилий и млекопи-
тающих) или Z-полосок (у амфибий) они образуют характерные комплексы, названные триадами. Под влиянием тока действия из цистерн саркоплазматической сети в гиалоплазму выходят ионы Са++, и когда их концентрация поднимается выше пороговой величины, Са включает механизм перемещения протофибрилл друг относительно друга. Сокращение идет с затратой энергии АТФ до тех пор, пока длится потенциал действия и в гиалоплазме достаточно много ионов Са++. Чем их больше, тем интенсивнее сокращаются миофибриллы. При исчезновении потенциала действия Са++ устремляются из гиалоплазмы в сарко-плазматическую сеть и мышечное волокно расслабляется.
Энергия для работы мышечного волокна образуется в процессе гликолиза и окислительного фосфорилирования. В зависимости от того, какой источник используется, в волокне будут преобладать включения гликогена или миоглобина. Волокна, содержащие большое количество миоглобина, имеют более красную окраску (в нефиксированной ткани) и названы красными. Волокна, в которых миоглобина меньше, имеют более светлый цвет и названы белыми. Между этими крайними типами есть промежуточные формы. Волокна отличаются друг от друга не только количеством миоглобина, но и структурно-функциональными особенностями. Как правило, каждая мышца содержит и белые, и красные, и промежуточные волокна. Ее свойства, хотя и зависят от того, какой тип волокон в ней преобладает, будут складываться не просто из суммы свойств составляющих ее волокон, но и зависеть от их взаимодействия в процессе работы.
Развитие поперечнополосатой скелетной мышечной ткани.
Данная ткань развивается из миотомов сомитов мезодермы. Миотомы состоят из плотно расположенных удлиненных клеток — миобластов. В период эмбриогенеза миобласты активно делятся, перемещаются в места закладки мускулатуры, выстраиваются в цепочки и сливаются друг с другом, образуя мышечные трубочки. В процессе дальнейшей дифференцировки происходит наращивание и совершенствование структур органелл и мышечные трубочки превращаются в мышечные волокна — дефинитивные симпла-
www.timacad.ru
Вракин В.Ф, Сидорова М.В. |
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ |
|
|
стические структуры поперечнополосатой скелетной мышечной ткани. Часть миобластов не проходит указанный путь, а сохраняется в виде удлиненных одноядерных малодифференцированных клеток, плотно прилежащих к волокну и одетых общей с ним базальной пластинкой. Такие клетки называют миосателлитами. Считают, что они могут быть источником образования новых волокон в течение всей жизни животного. Есть данные о том, что новые волокна образуются путем продольного расщепления толстых волокон и увеличения количества миофибрилл в тонких волокнах.
Возрастные и другие изменения скелетной мышечной ткани
выражаются прежде всего в увеличении размеров волокон. Диаметр волокна увеличивается за счет прибавления протофибрилл по бокам миофибрилл. В длину волокно растет в результате присоединения протофибрилл по концам миофибрилл. Увеличивается и количество миофибрилл в основном за счет продольного расщепления. Процессы эти активно протекают как в эмбриональный, так и в постнатальный период. За это время мышечные волокна утолщаются в 2—4 раза. У крупного рогатого скота особенно активно растут мышцы от рождения до 6 и от 12 до 18 месяцев. С возрастом у всех животных увеличивается относительное содержание в скелетных мышцах белых волокон по сравнению с красными.
Пучки мышечных волокон у новорожденных животных плохо сформированы, так как прослойки рыхлой соединительной ткани имеют вид нежной сети и не содержат жира. С ростом животного увеличивается количество пучков благодаря их разделению в результате врастания соединительной ткани. Мышечные пучки хорошо очерчены, лежат плотно. Прослойки перимизия тонкие, но более плотные, чем у новорожденных, в них больше становится коллагеновых волокон. Общее содержание соединительной ткани в мышцах колеблется в широких пределах (от 5 до 20%) в зависимости от типа мышцы. Старческие изменения в мышечной ткани выражаются в появлении большого количества ядер в центре волокон, разрастании плотной соединительной ткани, истончении и нарастании дегенеративных изменений в мышечных волокнах, снижении васкуляризации.
Мышечная ткань у животных разных пород и направления продуктивности может отличаться толщиной волокон, степенью развития и плотностью перимизия и эпимизия, хотя различия эти в основном незначительны. Все же замечено, что у животных скороспелых пород и мясного типа мышечные волокна более толстые: в их межпучковой соединительной ткани чаще встречаются жировые клетки. От кастрации замедляется рост волокон, сохраняется рыхлость перимизия. Стойловое содержание приводит к увеличению диаметра мышечных волокон и разрастанию жира в перимизии.
Значительно меняется структура мышечной ткани под влиянием кормления. Сбалансированное оптимальное кормление способствует нормальному развитию мышечной ткани. При обильном кормлении и откорме утолщаются мышечные волокна, между ними появляется много жировых клеток. Несбалансированное и неполноценное кормление несколько сдерживает рост
www.timacad.ru
Вракин В.Ф, Сидорова М.В. |
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ |
|
|
мышечной ткани. Пагубно сказывается на ней недостаточное питание, особенно в период наиболее интенсивного роста: диаметр мышечных волокон в этом случае меньше на 20—35%.
СЕРДЕЧНАЯ ПОПЕРЕЧНОПОЛОСАТАЯ (ИСЧЕРЧЕННАЯ) МЫШЕЧНАЯ ТКАНЬ
Сердечная ткань дифференцируется из висцерального листка спланхнотома мезодермы и образует основной слой стенки сердца— миокард. По структуре и функции различают две ее разновидности: рабочую и проводящую.
Рабочая мышечная ткань по структуре и физиологическим свойствам занимает как бы промежуточное положение между скелетной и гладкой. Сокращается она ритмично, медленнее скелетной мышечной ткани и мало утомляется. Состоит из клеток — сердечных миоцитов, объединенных в сердечные волокна. Последние анастомозируют друг с другом так, что образуют единую систему. Между волокнами имеются прослойки рыхлой соединительной ткани — эндомизий, в котором проходят сосуды и нервы.
Одеты сердечные волокна сарколеммой, подобной сарколемме скелетного мышечного волокна. Однако в отличие от него внутренний слой сарколеммы — плазмолемма — одевает каждую клетку отдельно. В местах соединения соседних клеток друг с другом их плазмолеммы образуют пальцевидные выпячивания и контакты типа десмосом, плотные и щелевые, а также специализированные контакты на уровне Z-полосок, которые способствуют функционированию сердечной мышцы как единого целого. Границы соседних миоцитов в волокнах представлены в виде вставочных (темных) полосок, идущих поперек волокна. С возрастом они утолщаются, особенно у крупного рогатого скота и лошадей.
Сердечные 1миоциты — клетки цилиндрической формы длиной 50—120 мкм и шириной 15—20 мкм. Однодва овальных ядра лежат в центре клетки. Миофибриллы занимают ее периферию. Строение их такое же, как в скелетной мышечной ткани, а количество меньшее. Поэтому поперечная исчерченность сердечных миоцитов видна хуже, чем продольная.
Сердечная мышечная ткань темно-красного цвета, так как цитоплазма сердечных миоцитов богата миоглобином. В ней много митохондрий и включений гликогена. Следовательно, она получает энергию как в процессе окислительного фосфорилирования, так и гликолиза. Благодаря мощной энергетической оснащенности сердечная мышца работает непрерывно всю жизнь. Замечено, что у животных, обитающих в высокогорных условиях, увеличивается количество гликогена, митохондрии крупнее, с волнистыми или сетчатыми кристами. Т-каналы в сердечных миоцитах расположены на уровне Z-полосок, а не на границе дисков, как в скелетных мышечных волокнах. Саркоплазматическая сеть развита хуже, поэтому сердечная мышца нуждается в постоянном притоке Са извне.
www.timacad.ru