

Стереоизомерия.
Все стандартные аминокислоты содержат
ассиметрический атом углерода в
-положении, т.е. атом
углерода с четырьмя различными
заместителями. Такой атом углерода
является хиральным центром. Благодаря
тому, что связи вокруг -атома
углерода имеют тераэдрическое
расположение, четыре различных заместителя
могут располагаться в пространстве
двумя различными способами, так что
молекула может существовать в двух
к онфигурациях,
представляющих собой несовместимые
зеркальные отображения.
онфигурациях,
представляющих собой несовместимые
зеркальные отображения.
Таким образом, соединения с хиральным центром встречаются в двух изомерных формах, у которых одинаковые физические и химические свойства, за исключением одного - способности вращать плоскость плоскополяризованного луча света в разные стороны на определенный угол. Эти соединения обладают оптической активностью. В основе строгой системы классификации и обозначения стереоизомеров лежит не вращение плоскости поляризации света, а абсолютная конфигурация молекулы стереоизомера, т.е. взаимное расположение четырех заместителей. Для выяснения конфигурации оптически активных соединений их сравнивают с каким-нибудь одним соединением, выбранным в качестве эталона, например, глицеральдегидом.

Почти все природные биологически активные соединения, содержащие хиральный центр, встречаются только в какой-нибудь одной стереоизомерной форме - D или L. Все аминокислоты, входящие в состав белков, являются L-изомерами. Живые клетки обладают уникальной способностью синтезировать L-аминокислоты с помощью стереоспецифичных ферментов. Стереоспецифичность этих ферментов обусловлена ассимитрическим характером их активных центров.

Кислотно-основные свойства аминокислот
Изоэл точка это значение рН, при которой концентрация биполярного иона в растворе максимальна, а концентрация аниона – минимальна (т.е. молекула заряжена нейтрально).
При растворении в воде кислоты ионизируются и ведут себя как кислоты и основания. Знание кислотно-основных свойств аминокислот имеет исключительно важное значение для понимания многих свойств белков.
По кислотно-основным свойствам аминокислоты разделяют на три группы.
Н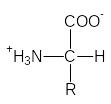 ейтральные
аминокислоты не содержат в радикале
R дополнительных кислотных
или основных центров, способных к
ионизации в водной среде. В кислой среде
они существуют в виде однозарядного
катиона и являются двухосновными
кислотами по Бренстеду. Как видно на
примере аланина, изоэлектрическая точка
у нейтральных аминокислот не равна 7, а
лежит в интервале 5,5 – 6,3.
ейтральные
аминокислоты не содержат в радикале
R дополнительных кислотных
или основных центров, способных к
ионизации в водной среде. В кислой среде
они существуют в виде однозарядного
катиона и являются двухосновными
кислотами по Бренстеду. Как видно на
примере аланина, изоэлектрическая точка
у нейтральных аминокислот не равна 7, а
лежит в интервале 5,5 – 6,3.
-Аминокислоты, содержащие одну аминогруппу и одну карбоксильную группу, кристаллизуются из нейтральных водных растворов в виде биполярных ионов (цвиттерионов).
Аминокислоты могут вести себя как кислоты (доноры протонов)
и как основания (акцепторы протонов):
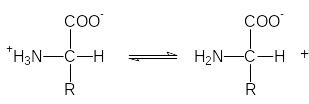
Д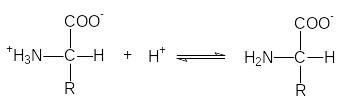 анные
равновесия можно количественно описать
константами диссоциации К1
и К2 или их отрицательными
десятичными логарифмами:
анные
равновесия можно количественно описать
константами диссоциации К1
и К2 или их отрицательными
десятичными логарифмами:
![]()
В зависимости от рН в растворе могут наблюдаться следующие равновесия:
p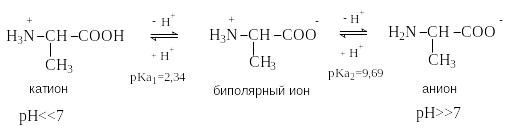 I=1/2(2,34+9,69)=6,01
I=1/2(2,34+9,69)=6,01
Основные аминокислоты содержат в радикале R дополнительный основный центр. К ним относятся лизин, гистидин и аргинин. В кислой среде они существуют в виде дикатиона и являются трехосновными кислотами. Изоэлектрическая точка основных аминокислот, как видно на примере лизина, лежит в области рН выше 7.
pI= ½(9,0+10,05)=9,74

Кислые аминокислоты содержат в радикале R дополнительный кислотный центр. К ним относятся аспаргиновая и глутаминовая кислоты. В кислой среде они существуют в виде катиона и являются трехосновными кислотами. Изоэлектрическая точка этих аминокислот лежит в области рН много ниже 7.
p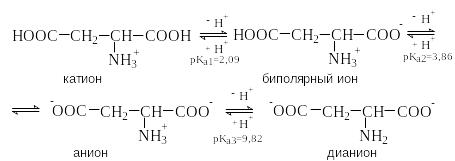 I=
½(2,09+3,86)=2,77
I=
½(2,09+3,86)=2,77
Тирозин и цистеин содержат в боковых радикалах слабые кислотные центры, способные к ионизации при высоких значениях рН.
Важное значение имеет тот факт, что при физиологическом значении рН (~7) ни одна аминокислота не находится в изоэлектрической точке. В организме все аминокислоты ионизированы, что обеспечивает им хорошую растворимость в воде.
Частота, с какой аминокислоты встречаются в белках, неодинакова. Например, глицин обнаруживается в 10 раз чаще, чем триптофан. По частоте нахождения аминокислот в белках можно составить такой ряд: ала вал лей сер глу глн лиз арг про > асп асн изо тре фен > тир цис мет гис.
Большинство белков по аминокислотному составу отличаются не очень резко. Но некоторые белки с особыми свойствами отличаются и аминокислотным составом. Так, белок соединительной ткани коллаген на 1/3 построен из остатков глицина, около 1/5 на ост. пролина и оксипролина. Именно такой состав аминокислот позволяет готовой молекуле белка образовывать прочные олигомерные структуры - фибриллы. Фибриллы коллагена превосходят по прочности стальную проволоку равного поперечного сечения. При кипячении в воде нерастворимый коллаген превращается в желатину - растворимую смесь полипептидов. Необычный аминокислотный состав коллагена определяет его низкую питательную ценность. В состав связок и соединительной ткани стенок сосуда входит белок - эластин. Эластин богат остатками лизина. Четыре боковые группы лизина сближаются друг с другом и ферментативным путем превращаются в десмозин. Таким путем полипептидные цепи эластина могут объединяться в системы, способные обратимо растягиваться во всех направлениях.
В хромосомах содержатся положительно заряженные белки гистоны, примерно на 1/3 построенные из остатков лизина и аргинина. Положительный заряд молекулы белка позволяет образовывать прочные комплексы с отрицательно заряженными молекулами нуклеиновых кислот.