

Список ферментов
Все известные ферменты занесены в специальный справочник, в котором информация о ферментах суммирована в виде таблицы.
Таблица. Список ферментов
Шифр |
Рабочее название |
Реакция |
Систематическое название |
Примечание |
КФ1.1.1.27 |
Лактатдегидрогеназа |
L-лактат + НАД+ пируват + НАДНН+ |
L-лактат:НАД+-оксидоредуктаза |
Окисляет и др. оксимонокарбоновые кислоты |
В настоящее время существуют постоянно пополняющиеся базы данных, в которых содержится информация обо всех известных ферментах. Эти базы данных доступны в Интернете. Например, адрес в Интернете международной базы данных "Enzyme nomenclature database": http://expasy.hcuge.ch/sprot/enzyme.html
Механизм действия ферментов. Активный центр ферментов
Ферменты (с точки зрения строения) – это глобулярные белки, которые имеют в глобуле полость или щель определенных размеров – активный центр фермента.
4.1.1 Общие черты активного центра фермента:
а.ц.ф. – небольшая часть белковой молекулы;
а.ц.ф. – имеет строго определенную трехмерную структуру и размеры;
а.ц.ф. – формируется из аминокислотных остатков, удаленных в первичной структуре, но сближенных в третичной структуре;
а.ц.ф. – состоит из связывающего и каталитического участков.
Если белок сложный, то простетическая группа входит в состав а.ц. фермента.
Вся белковая молекула укладывается определенным образом в пространстве, образуя единственно возможный (термодинамически устойчивый) вариант третичной структуры данного белка, а следовательно и уникальный активный центр данного фермента.
Механизм действия ферментов (на примере фермента холинэстеразы)
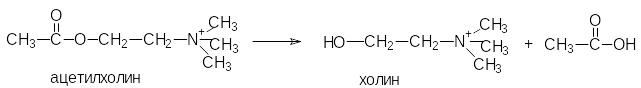
Субстрат данного фермента: ацетилхолин – медиатор передачи нервного импульса.
В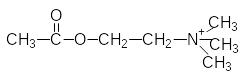
 ответ на выделение окончанием нервного
волокна ацетилхолина, следует ответная
реакция возбуждения нервной клетки.
Чтобы этот процесс протекал непрерывно,
после каждого акта передачи нервного
импульса вызвавшая возбуждение порция
ацетилхолина должна быть гидролизована.
Скорость гидролиза: 1-2 мкг (порция) за
0,1-0,2 мс.
ответ на выделение окончанием нервного
волокна ацетилхолина, следует ответная
реакция возбуждения нервной клетки.
Чтобы этот процесс протекал непрерывно,
после каждого акта передачи нервного
импульса вызвавшая возбуждение порция
ацетилхолина должна быть гидролизована.
Скорость гидролиза: 1-2 мкг (порция) за
0,1-0,2 мс.
Активный центр фермента состоит из двух функционально важных и пространственно разделенных участков:
связывающего, куда входит карбоксильная группа -COO- , электростатически взаимодействующая с заряженным азотом N+ субстрата;
каталитического, ответственного за эстеразную активность фермента, в состав которого входят остатки Ser, His,Tir.
В процессе реакции атом водорода гидроксильной группы Tir активного центра связывается с атомом кислорода ацетилхолина (будущая спиртовая группа продукта реакции – холина). В результате увеличивается положительный заряд на углеродном атоме ацетильной группы субстрата, который атакуется отрицательно заряженным атомом кислорода серина. Отрицательный заряд на атоме кислорода серина возникает в результате образования водородной связи между атомом Н серина и атомом N гистидина. Связь между С (ацетила) и О (холина) разрывается с образованием в качестве промежуточного соединения ацетилсерина. Отщепляющийся от серина протон связывается кислородным атомом тирозина, и первоначальное состояние тирозина восстанавливается. Гидролиз ацетилсерина начинается с диссоциации молекулы воды за счет взаимодействия протона с атомом N гистидина. Освободившийся гидроксил атакует сложноэфирную связь ацетилсерина. Результатом гидролиза является освобождение уксусной кислоты. Ион водорода (Н+), временно связанный с гистидином, освобождается и связывается с кислородом серина. Образовавшиеся холин и уксусная кислота освобождаются из активного центра за счет диффузии.
Все описанные выше процессы более или менее одновременно. Гидролиз ацетилхолина происходит благодаря согласованному действию всех функциональных групп активного центра.