

Пассивный транспорт
Возможны два типа пассивного транспорта:
Простая диффузия. Гидрофобная внутренняя часть мембраны является барьером для полярных молекул, но малые незаряженные или гидрофобные молекулы легко диффундируют через липидный бислой. Легче всего проходят простой диффузией через липидную мембрану малые неполярные молекулы, такие как О2, стероиды, тиреоидные гормоны, а также жирные кислоты. Малые полярные незаряженные молекулы - СО2, NH3, Н2О, этанол, мочевина - также диффундируют с достаточно большой скоростью. Диффузия глицерола идёт значительно медленнее, а глюкоза практически не способна самостоятельно пройти через мембрану. Для всех заряженных молекул, независимо от размера, липидная мембрана непроницаема.
Облегченная диффузия. Для переноса заряженных молекул функционируют специальные белки. Выделяют два основных класса белков-переносчиков:
каналообразующие белки, которые формируют в мембране поры, заполненные водой;
белки, связывающие переносимую молекулу с изменением конформации так, что переносимое вещество оказывается внутри клетки (или наоборот).
Такой тип переноса веществ по градиенту концентраций называется облегченной диффузией.
Белок-переносчик имеет центр связывания, комплементарный переносимому веществу, поэтому для облегченной диффузии, в отличие от простой, характерна высокая избирательность.
Активный транспорт
Клеткам необходимо перемещать вещества против градиента концентраций. Такое перемещение требует затрат энергии и называется активным транспортом. Как правило, белки-переносчики являются одновременно ферментами (АТФ-азами), способными гидролизовать АТФ. Таким образом, перенос веществ через мембрану против градиента концентраций сопряжен с гидролизом АТФ. 30-40% всей потребляемой энергии клетками расходуется на поддержание внутреннего содержания клетки за счет активного транспорта (Na+-K+-АТФаза).
В зависисмости от способа использования энергии для транспорта молекул выделяют первично активный транспорт и вторично активный транспорт.
При первично активном транспорте донором энергии является непосредственно молекула АТФ и процесс переноса вещества через мембрану сопровождается ее гидролизом.
При вторично активном транспорте градиент ионов, созданный функционированием систем первично активного транспорта, используется для переноса других молекул (углеводов, аминокислот и др.)
Для активного транспорта, как и для облеченной диффузии характерна высокая специфичность, эффект насыщения транспортных белков, а также действие ингибиторов.
Na+/K+-насос
В животной клетке концентрация ионов калия примерно в 30 раз выше, а ионов натрия в 10 раз ниже, чем в окружающей среде.
Na+/K+-насос – система первично-активного транспорта, отвечающая за поддержание высокой концентрации ионов калия и низкой концентрации ионов натрия внутри клетки путем переноса К+ внутрь клетки и Na+ наружу.
Перенос ионов натрия и калия осуществляет олигомерный. Интегрированный в мембрану белок, называемый Na+, К+-АТФ-аза.
Функционирование Na+/K+-насоса можно представить следующим образом:
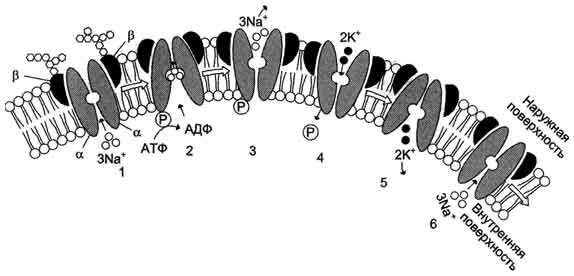
1 - три иона натрия связываются специфическим центром транслоказы.
2 - изменение конформации транслоказы, вызванное присоединением 3Na+, приводит к активации каталитической субъединицы и увеличению сродства активного центра к субстрату (АТФ). Протекает реакция аутофосфорилирования по карбоксильной группе аспарагиновой кислоты.
3 - аутофосфорилирование изменяет заряд и конформа-цию транслоказы, она закрывается с внутренней стороны мембраны и открывается с наружной, уменьшается сродство к ионам натрия и они диссоциируют от переносчика.
4 - Na+, К+-АТФ-аза открытая с наружной стороны мембраны имеет специфический центр связывания для 2К+. Присоединение двух ионов калия к фосфорилированной транслоказе вызывает изменение конформации и появление аутофосфатазной активности. Протекает реакция аутодефосфорилирования.
5 - дефосфорилирование изменяет заряд и конформацию транслоказы, она закрывается с наружной стороны мембраны и открывается с внутренней, уменьшается сродство к ионам калия и они диссоциируют от Na+, К+-АТФ-азы.
6 - АТФ-аза возвращается в первоначальное состояние.
Таким образом, на каждую затраченную молекулу АТФ происходит перенос трех ионов натрия из клетки и двух ионов калия в клетку.