

7. Обмен липидов.
7.1. Катаболизм глицерина.
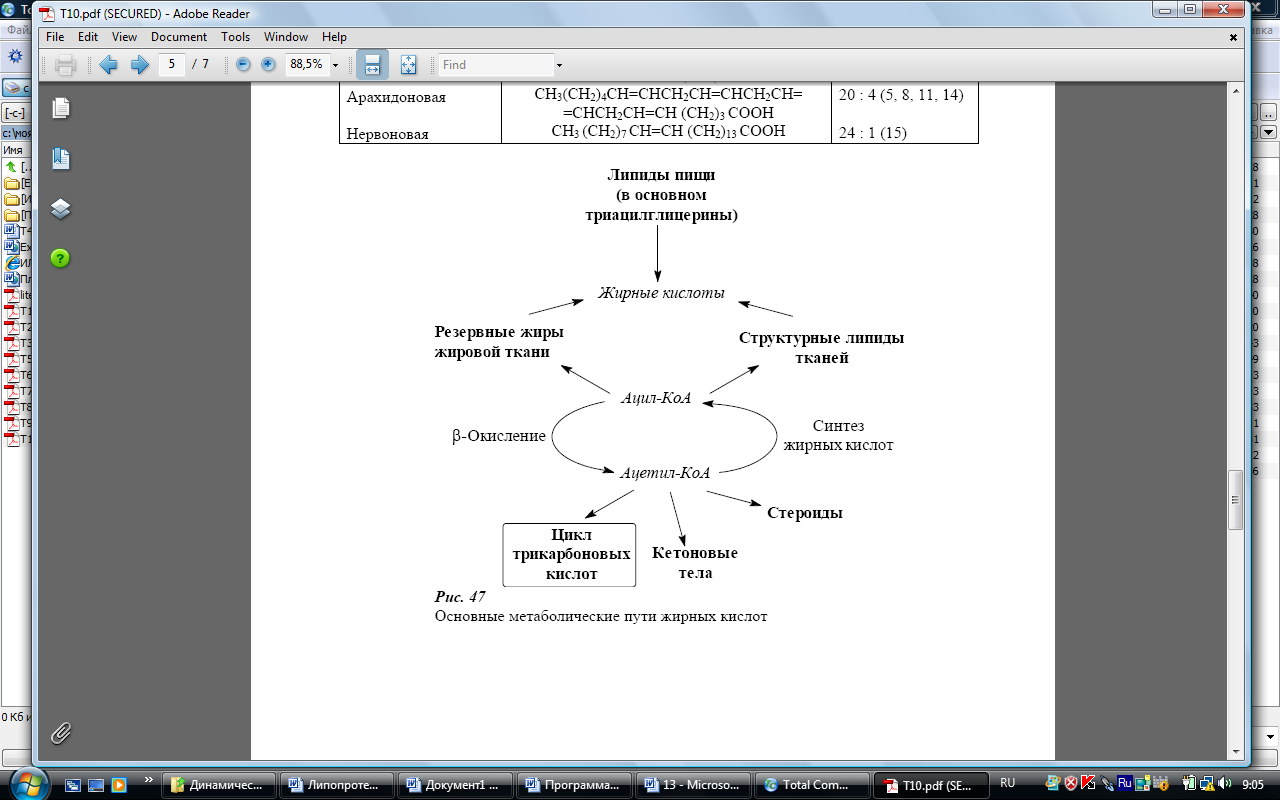
Под действием ферментов липаз триацилглицеролы в цитоплазме клетки гидролизуются до глицерола и свободных высших жирных кислот. Свободные жирные кислоты либо вновь вовлекаются в синтез липидов, либо подвергаются β-окислению, либо диффундируют в плазму крови и транспортируются в другие ткани.
Глицерол в цепи реакций может превращаться в глицеральдегид-3-фосфат и далее включаться в гликолиз или глюконеогенез.

7.2. -Окисление насыщенных и ненасыщенных жирных кислот.
При переваривании жиров образуется глицерин, жирные кислоты, диацилглицеролы и 75% моноацилглицеролов. Эта смесь продуктов гидролиза вместе с желчными кислотами, необходимыми для переваривания и всасывания липидов, поступает в кровь и с кровотоком - в печень. Жиры в организме запасаются в специализированных клетках жировой ткани – липоцитах. Главная роль жиров в организме – долговременный резерв энергии. Запаса жиров хватает на несколько недель голодания. Кроме того, жирные кислоты, входящие в состав жиров, участвуют в анаболических процессах (синтез структурных липидов тканей, холестерола). Жиры могут не только поступать с пищей, но и синтезироваться в организме из аминокислот и углеводов.
Окисление до со2 и образование атф
Свободные жирные кислоты подвергаются деградации с образованием ацетил-КоА в цитоплазме. Этот процесс называется -окислением жирных кислот. Ацетил-КоА далее окисляется в митохондриях в цикле лимонной кислоты. В результате полного окисления 1 моля пальмитиновой кислоты (С15Н33СООН) может образоваться 129 моль АТФ. Жирные кислоты служат основным субстратом энергетического обмена в печени.

Рис. Схема β-окисления жирных кислот.
7.3. Биосинтез липидов.
В настоящее время в достаточной степени изучен механизм биосинтеза жирных кислот в организме животных и человека, а также катализирующие этот процесс ферментные системы. Синтез жирных кислот протекает в цитоплазме клетки. В митохондриях в основном происходит удлинение существующих цепей жирных кислот. Установлено, что в цитоплазме печеночных клеток синтезируется пальмитиновая кислота (16 углеродных атомов), а в митохондриях этих клеток из уже синтезированной в цитоплазме клетки пальмитиновой кислоты или из жирных кислот экзогенного происхождения, т.е. поступающих из кишечника, образуются жирные кислоты, содержащие 18, 20 и 22 углеродных атома.
Иными словами, митохондриальная система биосинтеза жирных кислот, включающая несколько модифицированную последовательность реакций β-окисления, осуществляет только удлинение существующих в организме среднецепочечных жирных кислот, в то время как полный биосинтез пальмитиновой кислоты из ацетил-КоА активно протекает в цитозоле, т.е. вне митохондрий, по совершенно другому пути.
Образование малонил-КоА. Первой реакцией биосинтеза жирных кислот является карбоксилирование ацетил-КоА, для чего требуются бикарбонат, АТФ, ионы марганца. Катализирует эту реакцию фермент ацетил-КоА-карбоксилаза. Фермент содержит в качестве простетической группы биотин.
Энзиматические системы, осуществляющие синтез жирных кислот, называются жирно-кислотными синтетазами. Они широко встречаются в природе и могут быть изолированы из различных одноклеточных организмов, растений и животных тканей.
Жирно-кислотные синтетазы делятся на 2 группы. К первой группе относятся полиэнзимные, не поддающиеся фракционированию комплексы с мол. м. порядка 500000, в которых все индивидуальные энзимы собраны в компактную структуру. В частности, в эту группу входят жирно-кислотные синтетазы животных тканей и дрожжей.
Вторая группа включает жирно-кислотные синтетазы, из которых отдельные энзимы могут быть выделены методами белкового фракционирования. Такие синтетазы встречаются у ряда микроорганизмов (в частности, у E.coli) и растений. Иными словами, в этих случаях все индивидуальные ферменты синтетазной системы находятся в виде автономных полипептидов.
Мультиферментный комплекс, называемый синтетазой (синтазой) жирных кислот, состоит из 6 ферментов, связанных с так называемым ацилпереносящим белком (АПБ). Этот белок относительно термостабилен, имеет две свободные HS-группы (цистеина и фосфопантетеинового остатка, присоединенного к ОН-группе серина) и вовлекается в процесс синтеза высших жирных кислот практически на всех его этапах. Мол. масса АПБ составляет около 10000. Данный белок в синтетазной системе выполняет роль КоА. Заметим, что в животных тканях не удалось обнаружить свободного АПБ, подобного микробному. Из печени выделен полиэнзимный комплекс, содержащий все энзимы, необходимые для синтеза жирных кислот. Энзимы комплекса настолько прочно связаны друг с другом, что все попытки изолировать их в индивидуальном виде не увенчались успехом.
Последовательность реакций, происходящих при синтезе жирных кислот:
