

1.3. Особенности организации биосинтетических процессов в клетках прокариот и эукариот
Впервые серьёзное внимание на существенные отличия микроорганизмов от растений и животных обратил немецкий учёный Э. Геккель. Он предложил выделить все микроорганизмы в самостоятельное царство Протиста (Protista) наряду с царством растений (Plantae) и царством животных (Animalia). Подразделение протист на низшие и высшие произошло в соответствии с двумя выявленными типами клеточной организации – эукариотической и прокариотической. Высшие протисты – микроскопические животные (простейшие), микроскопические водоросли и микроскопические грибы (плесени и дрожжи), подобно растениям и животным, имеют эукариотическое строение клеток. Низшие протисты – бактерии, в том числе актиномицеты и синезелёные водоросли (цианобактерии), имеют прокариотическое строение.
Основные различия прокариот и эукариот:
Прокариоты могут существовать только в аэробных или только в анаэробных условиях, или и в тех и в других. Необходимую энергию они получают в процессе дыхания, брожения, фотосинтеза или хемосинтеза. Эукариоты – относятся к аэробным организмам.
У эукариот генетический материал локализован в структурно оформленном ядре. У прокариот наследственный материал представлен клубком двойной спиральной нити ДНК и не отделён от цитоплазмы какой – либо мембраной.
У прокариот, в отличие от эукариот, отсутствуют внутриклеточные органеллы, имеющие хотя бы элементарную мембрану.
У прокариот рибосомы имеют меньший размер.
Клеточная стенка прокариот содержит специальный полимер пептидогликан, которого нет у эукариот.
Жгутики прокариот состоят из одной или нескольких фибрилл, а у эукариот каждый жгутик состоит из микротрубочек – 9 по периметру и 2 в центре.
Многие прокариоты способны фиксировать молекулярный азот, эукариоты этим свойством не обладают.
Многие прокариоты способны выделять в окружающую среду продукты метаболизма (органические кислоты, витамины, антибиотики и др.)
2. Аминокислоты и белки
2.1. -Аминокислоты как структурные компоненты белков. Классификация аминокислот по характеру боковых заместителей. Кислотно-основные свойства аминокислот
Ключ к пониманию структуры любого белка дает небольшая группа довольно простых молекул - -аминокислот, играющих роль строительных блоков. Для построения всех белков используется один и тот же набор 20 различных, ковалентно связанных друг с другом в определенной, характерной только для данного белка последовательности. Каждая аминокислота благодаря особенностям ее боковой цепи наделена химической индивидуальностью, поэтому всю группу 20 аминокислот можно рассматривать как алфавит "языка" белковой структуры.
Строение и классификация аминокислот
В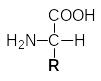 се
20 аминокислот, встречающихся в белках,
характеризуются общей структурной
особенностью - наличием карбоксильной
и аминогруппы, связанных с одним и тем
же атомом углерода. Различаются же
аминокислоты боковыми цепями (R-группами).
се
20 аминокислот, встречающихся в белках,
характеризуются общей структурной
особенностью - наличием карбоксильной
и аминогруппы, связанных с одним и тем
же атомом углерода. Различаются же
аминокислоты боковыми цепями (R-группами).
Формулы и тривиальные названия важнейших аминокислот приведены в таблице. Для биологического функционирования аминокислот в составе белков определяющим является полярность радикала R. По этому признаку аминокислоты разделяют на следующие основные группы.
Таблица. Важнейшие -аминокислоты RCH(NH2)COOH
Формула |
Название |
Обозначение |
pI |
Аминокислоты, содержащие неполярный радикал R
|
|||
|
Глицин |
Gly |
5,97 |
|
Аланин |
Ala |
6,0 |
|
Валин |
Val |
5,96 |
|
Лейцин |
Leu |
5,98 |
|
Изолейцин |
Ile |
6,02 |
|
Фенилаланин |
Phe |
5,48 |
|
Триптофан |
Trp |
5,89 |
|
Пролин (изгиб пептидной цепи) |
Pro |
6,30 |
|
Метионин |
Met |
5,74 |
|
Цистин |
(Cys)2 |
5,0 |
Аминокислоты, содержащие полярный неионогенный радикал R
|
|||
|
Серин |
Ser |
5,68 |
|
Треонин
|
Thr |
5,60 |
|
Гидроксипролин Не имеет генетический код |
Hyp |
5,8 |
|
Аспаргин |
Asn |
5,41 |
|
Глутамин |
Gln |
5,65 |
Аминокислоты, содержащие полярный положительно заряженный радикал R |
|||
|
Лизин |
Lys |
9,74 |
|
5-Гидроксилизин Не имеет генетич код |
|
9,15 |
|
Аргинин |
Arg |
10,76 |
|
Гистидин |
His |
7,59 |
Аминокислоты, содержащие полярный отрицательно заряженный радикал R |
|||
|
Аспаргиновая кислота |
Asp |
2,77 |
|
Глутаминовая кислота |
Glu |
3,22 |
|
Тирозин |
Tyr |
5,66 |
|
Цистеин |
Cys |
5,07 |








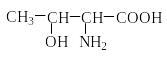


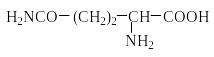

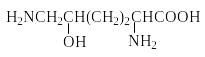






Белки-гистоны содержат много лизина и аргинина.
Аминокислоты, содержащие неполярный радикал R. Такие группы располагаются внутри молекулы белка и обуславливают гидрофобные взаимодействия.
Аминокислоты, содержащие полярный неионогенный радикал R. Аминокислоты этого типа имеют в составе бокового радикала полярные группы, не способные к ионизации в водной среде (спиртовый гидроксил, амидная группа). Такие группы могут располагаться как внутри, так и на поверхности молекулы белка. Они участвуют в образовании водородных связей с другими полярными группами.

Аминокислоты, содержащие радикал R, способный к ионизации в водной среде с образованием положительно или отрицательно запряженных групп. Такие аминокислоты содержат в боковом радикале дополнительный основный или кислотный центр, который в водном растворе может соответственно присоединять или отдавать протон.
В белках ионогенные группы этих аминокислот располагаются, как правило, на поверхности молекулы и обуславливают электростатические взаимодействия.