

Экзо- и эндоцитоз
Макромолекулы (белки, полисахариды и полинуклеотиды) и крупные частицы могут поглощаться и секретироваться клетками вместе с участком мембраны.
При процессе эндоцитоза поглощенное вещество окружается небольшим участком мембраны, который вначале выпячивается, а затем отщепляется, образую внутриклеточный пузырек, содержащий захваченный клекой материал.
Подобный процесс в обратном направлении называется экзоцитоз. Если таким путем осуществляется транспорт растворенных веществ, то это – пиноцитоз, твердых частиц – фагоцитоз.
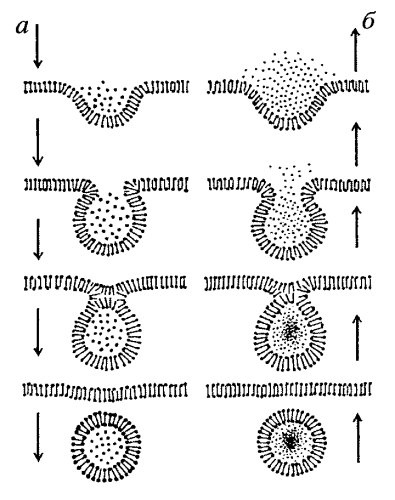
Рис. Эндо- (а) и экзоцитоз (б).
В изменении формы мембраны принимают участие сократительные белки, функционирование которых требует затрты АТФ. Поэтому процесс эндо- и экзоцитоза можно отнести к механизму активного транспорта.
Системы унипорта, симпорта и антипорта
Унипорт – независимый перенос одного вещества в одну сторону.
Симпорт – взаимосвязанный перенос двух веществ в одну строну.
Антипорт – взаимосвязанный перенос двух веществ в разные стороны через мембрану.

Рис. Типы облегчённой диффузии с участием переносчиков (транслоказ). S1, S2- разные молекулы.
4.3. Строение клеточной стенки грамположительных и грамотрицательных бактерий.
Бактерии можно разделить на два класса – грамположительные и грамотрицательные – в зависимости от их способности окрашиваться по методу, описанному в 1884 г. датским бактериологом Граммом. В процессе отмывки, которая следует за процедурой окрашивания, грамположительные бактерии удерживают фиолетовый краситель, а грамотрицательные теряют его. Это обусловлено различием в строении клеточных стенок двух указанных классов бактерий.
Установлено, что грамположительные бактерии обладают однослойной нелипидной клеточной стенкой толщиной 20-80 нм, окружающей плазматическую мембрану толщиной 7,5 нм. У грамотрицательных бактерий клеточная стенка двухслойная: первый слой – очень тонкая (1 нм) нелипидная мембрана, второй – липидная мембрана 7,5 нм.

Рис. Схема строения клеточной стенки грамположительных (а) и грамотрицательных (б) бактерий.
1 – плазматическая мембрана; 2 – слой пептидогликана; 3 – липопротеидный слой; 4 – периплазматическое пространство
Клеточная стенка – это жесткая структура, служащая для поддержания формы клетки и для предотвращения ее лизиса от осмотического шока. Материал клеточных стенок – пептидогликан (гликопептид, мукопептид).
Пептидогликан – сложный каркас, содержащий полисахаридные цепи, связанные сшивками из коротких пептидных цепей. Основа структуры пептидогликана – цепь параллельных полисахаридных цепей, построенных из чередующихся моносахаридов – N-ацетилглюкозамина (NAG) и N-ацетилмурамовой кислоты (NAM), связанных β(1→4)-гликозидными связями (до нескольких сотен моносахаридных блоков). Эти цепи образуют трехмерную волокнистую структуру до 40 слоев у грамположительных бактерий и один слой грамотрицательных.

Рис. Фрагмент полисахаридной цепи пептидогликана, построенной из чередующихся моносахаридов – N-ацетилглюкозамина (NAG) и N-ацетилмурамовой кислоты (NAM), соединенных β(1→4)-гликозидными связями.
Поперечные сшивки содержат два типа структур:
Тетрапептидный элемент, который присоединен к карбоксильной группе NAM при С3.
Пептидный мостик обычно представляет собой пентапептидную цепочку, состоящую только из аминокислот с короткими боковыми цепями, наиболее распространенный – пентаглицин.
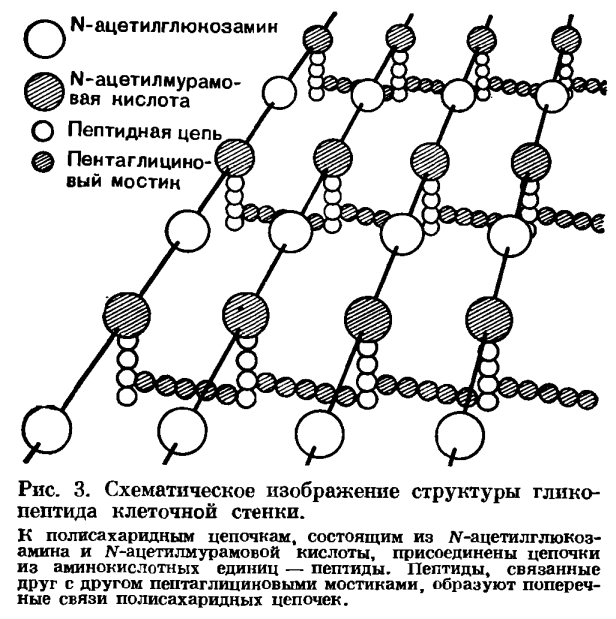
Рис. Схематическое изображение структуры пептидогликана.
Клеточную стенку грамположительных бактерий пронизывают тейхоевые и липотейхоевые кислоты, расположенные перпендикулярно плазматической мембране. Тейхоевые кислоты – это цепочки из молекул глицерола или рибитола, связанных фосфодиэфирными мостиками (до 30 молекул). В цепочку могут быть включены остатки сахаров и аминокислот. Таким образом, благодаря наличию тейхоевых кислот слои пепетидогликана оказываются связанными друг с другом.
