

5.4. Фотосинтез и хемосинтез. Фотосинтез
Первичным источником энергии на Земле является энергия солнца. Фотосинтезирующие организмы (растения, красные и бурые водоросли, цианобактерии, зеленые и пурпурные бактерии) обладают способностью улавливать кванты солнечного света и трансформировать их в энергию химических связей органических соединений.
![]()
Образующиеся в процессе фотосинтеза углеводы используются нефотосинтзирующими организмами как источник энергии и углерода. Кроме того, фотосинтез – единственный источник кислорода на Земле.
Таким образом, фотосинтез – это синтез углеводов и других органических соединений из неорганических (углекислого газа и воды) под действием энергии солнечного света.
Фотосинтез у растений протекает в клеточных органеллах – хлоропластах. Хлоропласты имеют форму двояковыпуклой линзы, размер их около 4-6 мкм. Находятся они в паренхимных клетках листьев и других зеленых частей высших растений. Число их в клетке варьирует в пределах 25-50. Снаружи хлоропласт покрыт оболочкой, состоящей из двух липопротеиновых мембран. Под ней, в основном веществе (строме), упорядоченно расположены многочисленные образования — ламеллы. Они образуют плоские мешочки – тилакоиды, которые лежат друг на друге правильными стопками. Эти стопки, напоминающие монеты, сложенные столбиком, называются гранами. Сквозь них проходят более длинные ламеллы, так что все граны хлоропласта связаны в единую систему. В состав мембран, образующих граны, входит зеленый пигмент — хлорофилл. Именно здесь происходят световые реакции фотосинтеза — поглощение хлорофиллом световых лучей и превращение энергии света в энергию возбужденных электронов.
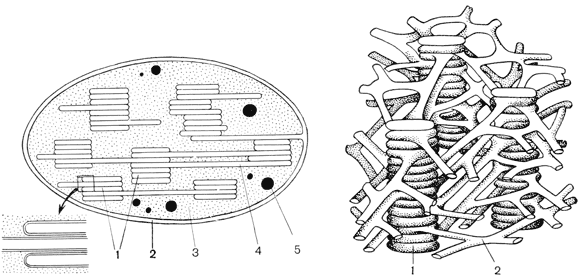
Рис. Строение хлоропласта. Слева — продольный разрез через хлоропласт. Участок внизу показан в увеличенном виде: 1 — граны, образованные ламеллами, сложенными стопками; 2 — оболочка; 3 — строма (матрикс); 4 — ламеллы; 5 — капли жира, образовавшегося в хлоропласте. Справа — трехмерная схема расположения и взаимосвязи ламелл и гран внутри хлоропласта: 1 — граны; 2 — ламеллы.
Фотосинтез пространственно и во времени разделяется на два сравнительно обособленных процесса: световую стадию окисления воды и темновую стадию восстановления CO2. Обе эти стадии осуществляются у высших растений и водорослей в хлоропластах. Исключение - синезеленые водоросли (цианобактерии), у которых нет аппарата фотосинтеза, обособленного от цитоплазматических мембран.
Световая стадия реализуется в мембранных структурах хлоропласта – тилакоидах, тогда как темновая стадия происходит в жидком содержимом хлоропласта (строме) при участии водорастворимых ферментов. У фотосинтезирующих бактерий хлоропласты отсутствуют, но световая стадия также осуществляется в мембранных образованиях - в так называемых хроматофорах.
Световая стадия фотосинтеза
Минимальная функциональная единица, способная осуществлять световую стадию фотосинтеза – тилакоид. Он представляет собой микроскопический плоский диск, образованный белковолипидными мембранами, в которых находятся пигменты. В эти мембраны встроены все компоненты, необходимые для окисления воды, восстановления кофермента никотинамиддинуклеотидфосфата (НАДФ) до НАДФН и синтеза АТФ из аденозиндифосфата. Световая стадия инициируется поглощением кванта света пигментами, организованными в специальные светособирающие комплексы. Среди пигментов преобладает хлорофилл а. К вспомогательным пигментам относятся хлорофилл b, каротиноиды и др.
Длина волны поглощаемого света зависит от структуры хлорофилла и его микроокружения в липопротеиновом слое мембраны тилакоидов.
Характерной особенностью химической структуры молекулы хлорофилла является наличие в порфириновом ядре замкнутой системы двойных сопряженных связей. Это обусловливает важнейшую физико-химическую особенность хлорофилла, имеющую биологическое значение – способность поглощать энергию света в видимом диапазоне электромагнитного спектра и преобразовывать эту энергию в энергию электронного возбуждения.
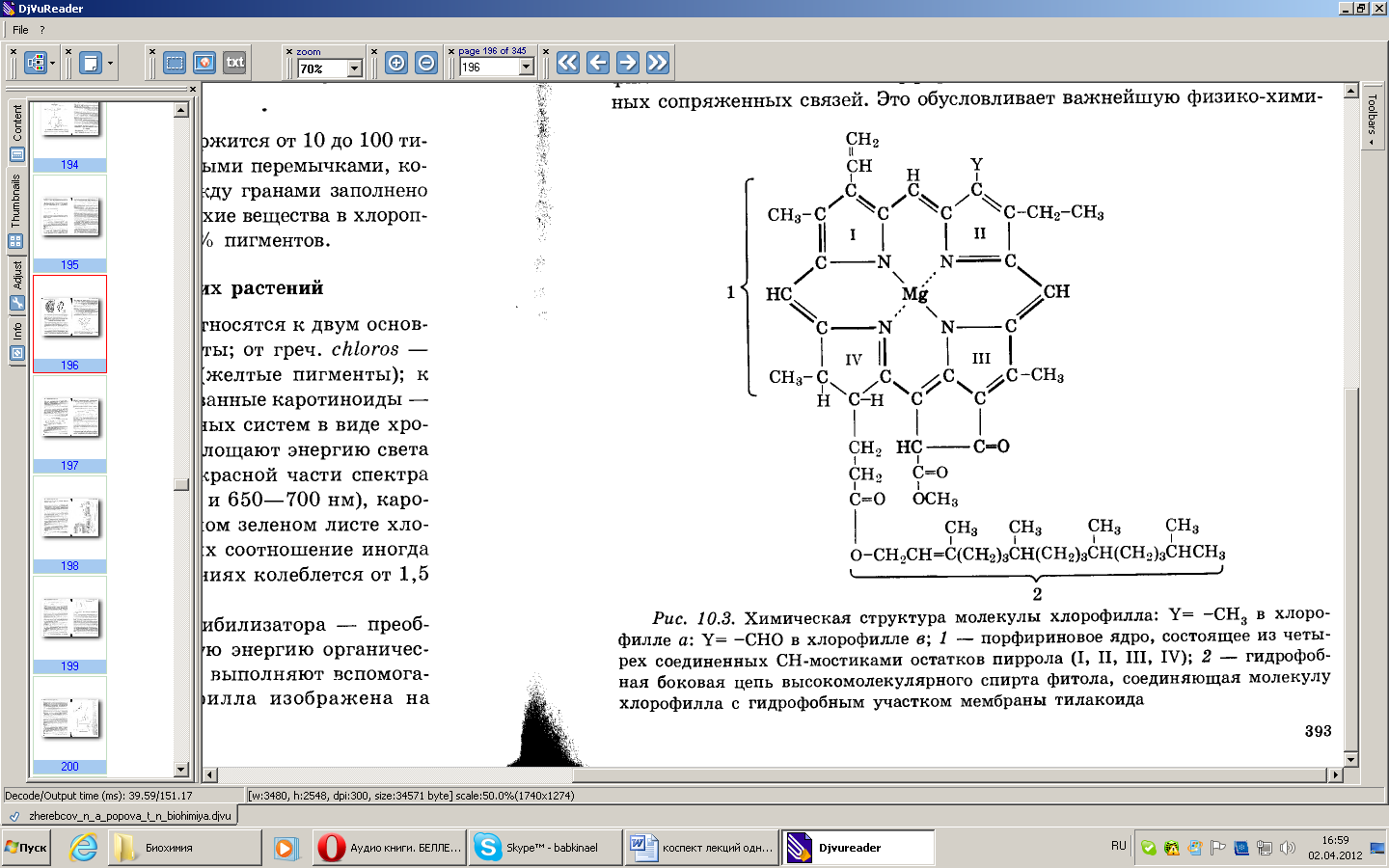
Рис. Химическая структура молекулы хлорофилла: Y= –CH3 в хлорофилле а; Y= –CHО в хлорофилле b; 1 – порфириновое ядро, состоящее из трех, соединенных СН-мостиками остатков пиррола (I, II, III, VI); 2 – гидрофобная боковая цепь высокомолекулярного спирта фитола, соединяющая молекулу хлорофилла с гидрофобным участком мембраны тилакоида.
Молекулы хлорофилла в хлоропластах организованы в крупные агрегаты – светособирающие системы. Часть вспомогательных пигментов, спектрально наиболее близких к фотохимически активному хлорофиллу, непосредственно окружает каждый из реакционных центров, образуя так называемые антенны.
Наличие такой светособирающей структуры из нескольких сотен или десятков молекул пигментов на каждый фотохимически активный (реакционный) центр на 2-3 порядка увеличивает сечение захвата излучения и обеспечивает возможность фотосинтеза при слабом освещении.