

Механизм сопряжения окисления и фосфорилирования
Свободная энергия, доступная в результате переноса двух электронов от НАДН и сукцината соответственно составляет 238 и 150 кДж/моль соответственно. На синтез одного моль АТФ расходуется 31 кДж/моль энергии. Синтез АТФ из АДФ сопряженный с процессом переноса электронов по дыхательной цепи и катализируемый интегральным белком - АТФ-синтазой, называется окислительным фосфорилированием. Суммарный процесс можно представить в следующем виде:
АДФ + Фн + энергия АТФ + Н2О
Сопряжение происходит при участии внутренней мембраны митохондрий.
Наибольшее признание получила хемиосмотическая теория сопряжения окисления и фосфорилирования, предложенная Митчеллом в 1961 г: протоны, перенесённые из матрикса в межмембранное пространство, не могут вернуться обратно в матрикс, так как внутренняя мембрана непроницаема для протонов. Таким образом, создаётся протонный градиент, при котором концентрация протонов в межмембранном пространстве больше, а рН меньше, чем в матриксе. Кроме того, каждый протон несёт положительный заряд, и вследствие этого появляется разность потенциалов по обе стороны мембраны: отрицательный заряд на внутренней стороне и положительный - на внешней. В совокупности электрический и концентрационный градиенты составляют электрохимический потенциал ΔμН+ - источник энергии для синтеза АТФ. Так как наиболее активный транспорт протонов в межмембранное пространство, необходимый для образования ΔμН+, происходит на участках ЦПЭ, соответствующих расположению комплексов I, III и IV, эти участки называют пунктами сопряжения дыхания и фосфорилирования.
Строение атф-синтазы и синтез атф
АТФ-синтаза (Н+-АТФ-аза) - интегральный белок внутренней мембраны митохондрий. Он расположен в непосредственной близости к дыхательной цепи. АТФ-синтаза состоит из 2 белковых комплексов, обозначаемых как F0 и F1.
Гидрофобный комплекс F0 погружён в мембрану. Он служит основанием, которое фиксирует АТФ-синтазу в мембране. Комплекс F0 состоит из нескольких субъединиц, образующих канал, по которому протоны переносятся в матрикс.
Комплекс F1 выступает в митоховдриальный матрикс. Он состоит из 9 субъединиц (3α, 3β, γ, ε, δ). Субъединицы α и β уложены попарно, образуя "головку"; между α- и β-субъединицами располагаются 3 активных центра, в которых происходит синтез АТФ; γ-, ε-, δ- субъединицы связывают комплекс F1 с F0.

Рис. Строение АТФ-синтазы.
Повышение концентрации протонов в межмембранном пространстве активирует АТФ-синтазу. Электрохимический потенциал ΔμH+ заставляет протоны двигаться по каналу АТФ-синтазы в матрикс. Параллельно под действием ΔμH+ происходят конформационные изменения в парах α, β-субъединиц белка F1, в результате чего из АДФ и неорганического фосфата образуется АТФ. Электрохимический потенциал, генерируемый в каждом из 3 пунктов сопряжения в ЦПЭ, используется для синтеза одной молекулы АТФ.
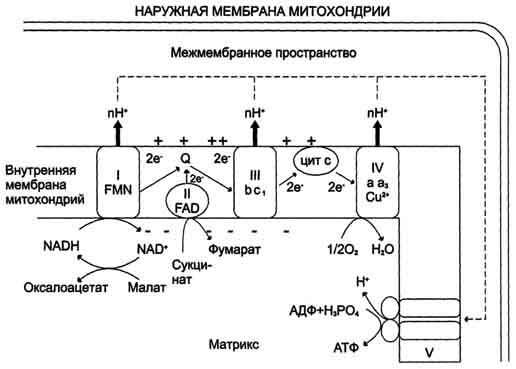
Рис. Сопряжение дыхания и синтеза АТФ в митохондриях. I - НАДН-дегидрогеназа; II - сукцинат дегидрогеназа; III - QН2-дегидрогеназа; IV - цитохромоксидаза; V - АТФ-синтаза. Энергия протонного потенциала (электрохимического потенциала ΔμН+ используется для синтеза АТФ, если протоны возвращаются в матрикс через ионные каналы АТФ-синтазы.