

8. Обмен белков
8.1. Общая схема метаболизма аминокислот. Механизмы транспорта аминокислот в клетку. Метаболизм аминокислот в печени
Б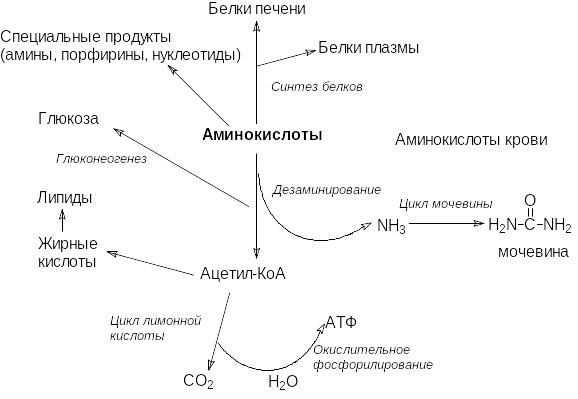 елки
перевариваются в желудочно-кишечном
тракте до аминокислот. Аминокислоты,
всосавшиеся в кишечнике и поступившие
затем в печень, также имеют несколько
путей метаболизма. Аминокислоты являются
важнейшими субстратами метаболизма
азота. От аминокислот берут начало
белки, ферменты, пуриновые и пиримидиновые
основания (и нуклеиновые кислоты),
пиррольные производные (порфирины),
биологически активные соединения
пептидной природы (гормоны), а также ряд
других соединений.
елки
перевариваются в желудочно-кишечном
тракте до аминокислот. Аминокислоты,
всосавшиеся в кишечнике и поступившие
затем в печень, также имеют несколько
путей метаболизма. Аминокислоты являются
важнейшими субстратами метаболизма
азота. От аминокислот берут начало
белки, ферменты, пуриновые и пиримидиновые
основания (и нуклеиновые кислоты),
пиррольные производные (порфирины),
биологически активные соединения
пептидной природы (гормоны), а также ряд
других соединений.
Транспорт в другие ткани
Аминокислоты из печени могут поступать в систему кровообращения и таким образом доставляться в другие органы и ткани, а там использоваться в качестве строительных блоков для биосинтеза тканевых белков.
Биосинтез белков печени и плазмы крови
Белки печени подвергаются постоянному обновлению, причем для них характерна очень высокая скорость оборота со средним периодом полужизни всего несколько дней. Кроме того, именно в печени синтезируется большинство белков плазмы крови.
Дезаминирование и распад
Участие в цикле глюкоза-аланин
Спустя несколько часов после приема пищи из мышц в печень поступает аланин, который подвергается деаминированию, а образующийсяпируват превращается в глюкозу. Дефицит аминокислот в мышцах пополняется после еды.
Превращение в нуклеотиды и другие продукты
Аминокислоты служат предшественниками в синтезе пуриновых и пиримидиновых оснований нуклеотидов, а также в синтезе некоторых специализированных веществ.
Окситоцин и вазопрессин – циклические пептиды гормонального действия.
Глутатион – трипептид, участвующий в системе обезвреживания ксенобиотиков.
Серотонин, гистамин – биогенные амины.
Меланин – пигмент кожи.
Адреналин – гормон.
Холин – предшественник нейромедиатора и фосфолипидов.
Креатин – предшественник креатинфосфата.
Порфирины.
Коферменты.
8.2. Дезаминирование аминокислот: окислительное дезаминирование, трансаминирование, непрямое дезаминирование (трансдезаминирование).
Те аминокислоты, которые не были использованы в печени или других органах для биосинтеза белков, подвергаются дезаминированию и с образованием ацетил-КоА и промежуточных субстратов цикла лимонной кислоты.
Чаще всего удаление
аминогруппы достигается переаминированием,
в ходе которого аминогруппа аминокислоты
переносится на -кетоглутаровую
кислоту, которая в результате превращается
в глутаминовую кислоту. Затем специфическими
дегидрогеназами глутаминовая кислота
дезаминируется с о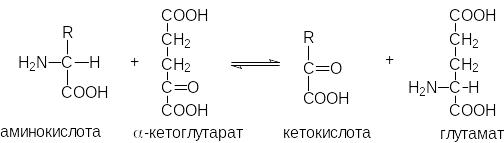 бразованием
на -кетоглутаровой
кислоты и аммиака.
бразованием
на -кетоглутаровой
кислоты и аммиака.
П![]() ереаминирование
аминокислот является важным связующим
звеном между метаболизмом аминокислот
(белков) и сахаров. В этот процесс
вовлечены заменимые гликогенные
аминокислоты, которые превращаются в
гликоген за счет глюконеогенеза через
ряд промежуточных продуктов цикла
лимонной кислоты.
ереаминирование
аминокислот является важным связующим
звеном между метаболизмом аминокислот
(белков) и сахаров. В этот процесс
вовлечены заменимые гликогенные
аминокислоты, которые превращаются в
гликоген за счет глюконеогенеза через
ряд промежуточных продуктов цикла
лимонной кислоты.
Углеродные атомы аминокислот могут включаться в общий путь катаболизма шестью путями:
А) из глицина, серина, цистеина, аланина, треонина образуется пируват (гликогенные аминокислоты)
Б) из лизина, тирозина, фенилаланина, лейцмна и триптофана образуется ацетил-КоА через ацетоацетил-КоА (кетогенные аминокислоты)
В) глутамин, глутамат, гистидин, пролин, аргинин вкючаются через -кетоглутарат
Г) четыре углеродных атома фенилаланина и тирозина включаются через фумарат
Д) метионин, изолейцин и валин включаются через сукцинил-КоА
Е) аспартат и асрарагин образуют оксалоацетат.
Аммиак, образующийся при дезаминировании, токсичен для организма и выводится в виде различных соединений. У человека конечным продуктом является мочевина. Мочевина образуется в результате процесса, который получил название цикла мочевины. Для биосинтеза мочевины требуется две молекулы аммиака. Источником аммиака первой реакции служит окислительное дезаминирование глутаминовой кислоты, для второй испрользуется аммиак аспарагиновой кислоты, которая образуется из глутаминовой путем переаминирования с оксалоацетатом.