

Макроэргические соединения
Энергия, освобождающаяся при реакциях гидролиза разных веществ, обычно невелика. Если она превышает 30 кДж/моль, то такую связь называют высокоэнергетической. Энергия гидролиза некоторых субстратов приведена в таблице.
Макроэргические соединения можно разделить на две группы:
Соединения, образующиеся в ходе процессов катаболизма и служащие для превращения энергии в качестве передаточного звена (АТФ; 1,3-дифосфоглицерат и фосфоенолпируват – промежуточные метаболиты катаболизма углеводов).
Соединения, которые могут быть использованы тканями в качестве "аварийного" источника энергии (креатинфосфат локализован в основном в мышцах и служит дополнительным источником энергии при стремительных кратковременных нагрузках).
Соединение |
-G', кДж/моль |
Фосфоенолпируват |
67,9 |
1,3-Дифосфоглицерат (продукт: 3-фосфоглицерат) |
49,3 |
Креатинфосфат |
43,1 |
АТФ АДФ + Ф |
30,5 |
АДФ АМФ + Ф |
27,6 |
Пирофосфат |
27,6 |
Глюкозо-1-фосфат |
20,9 |
АМФ |
14,2 |
Среднее положение АТФ по величине стандартной свободной энергии гидролиза позволяет ему служить донором высокоэнергетического фосфата для соединений, имеющих меньшую, чем АТФ величину стандартной свободной энергии гидролиза. В то же время при наличии соответствующих ферментных систем АДФ может акцептировать высокоэнергетический фосфат (с образованием АТФ) от тех соединений, которые обладают большей величиной стандартной свободной энергии гидролиза. Т.о., цикл АДФ/АТФ связывает процессы, генерирующие высокоэнергетический фосфат, с процессами, потребляющими энергию высокоэнергетических фосфатов.
Фазы освобождения энергии из питательных веществ
Характер метаболизма в тканях во многом определяется питанием. У человека метаболическим превращениям подвергаются, главным образом, глюкоза, жиры и аминокислоты. Глюкоза и аминокислоты - структурные компоненты белков и углеводов. Таким образом, необходимыми компонентами пищи являются белки углеводы и жиры.
В ходе извлечения энергии из различных субстратов можно условно выделить три фазы катаболизма питательных веществ.
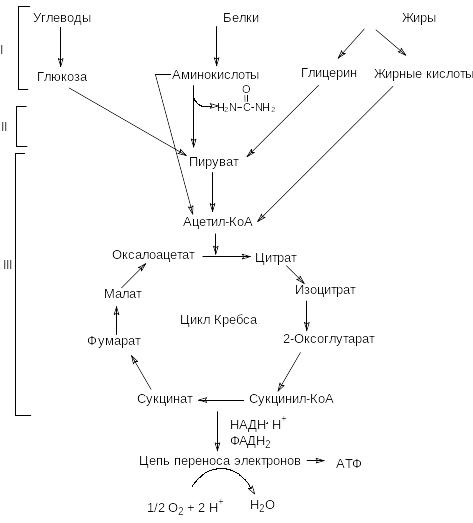
Рис. Фазы катаболизма белков, жиров, углеводов
Первая фаза - подготовительная. Она необходима для превращения полимеров, поступающих с пищей или находящихся внутри клетки, в удобную для извлечения энергии форму - мономеры. На этом этапе освобождается около 1% всей энергии, запасенной в питательных веществах. Эта энергия рассеивается в форме теплоты.
Вторая фаза - частичный распад мономеров до ключевых промежуточных продуктов: пирувата, ацетил-КоА и некоторых метаболитов цикла Кребса. На этом этапе освобождается около 20% энергии. Эта стадия называется промежуточный обмен. Реакции промежуточного обмена протекают в цитоплазме.
Третья фаза - окончательный распад всех веществ до CO2 и H2O. Этот этап включает реакции общего пути катаболизма (окислительное декарбоксилирование пирувата, цикл Кребса) и цепь переноса электронов. Примерно 80% всей энергии химических связей освобождается в данной фазе. Все реакции этой фазы локализуются в митохондриях.